

Drosophila ORB protein in two mushroom body output neurons is necessary for long-term memory formation
- PMID: 23610406
- PMCID: PMC3651462
- DOI: 10.1073/pnas.1216336110
Drosophila ORB protein in two mushroom body output neurons is necessary for long-term memory formation
Abstract
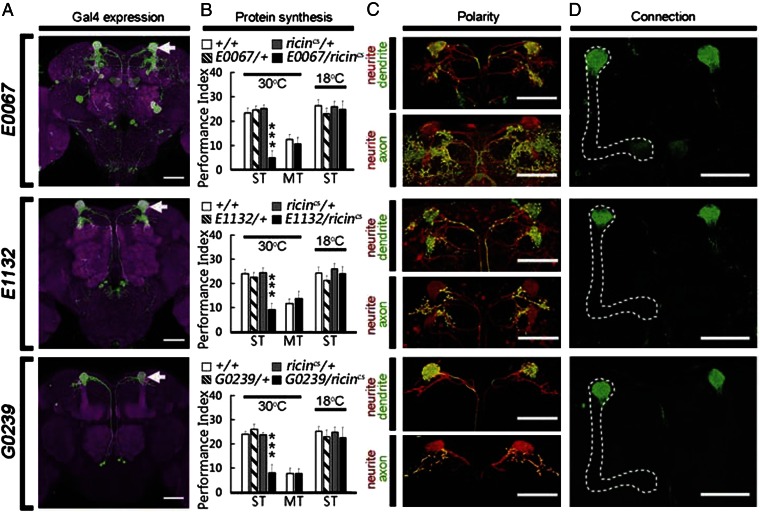
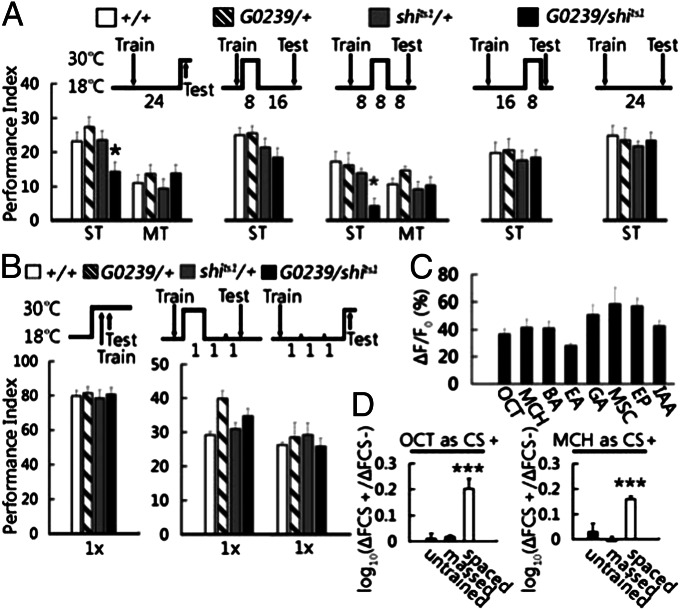
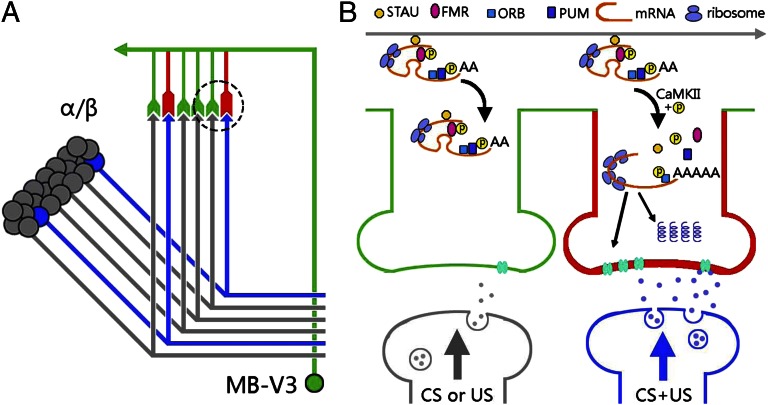
Memory is initially labile and gradually consolidated over time through new protein synthesis into a long-lasting stable form. Studies of odor-shock associative learning in Drosophila have established the mushroom body (MB) as a key brain structure involved in olfactory long-term memory (LTM) formation. Exactly how early neural activity encoded in thousands of MB neurons is consolidated into protein-synthesis-dependent LTM remains unclear. Here, several independent lines of evidence indicate that changes in two MB vertical lobe V3 (MB-V3) extrinsic neurons are required and contribute to an extended neural network involved in olfactory LTM: (i) inhibiting protein synthesis in MB-V3 neurons impairs LTM; (ii) MB-V3 neurons show enhanced neural activity after spaced but not massed training; (iii) MB-V3 dendrites, synapsing with hundreds of MB α/β neurons, exhibit dramatic structural plasticity after removal of olfactory inputs; (iv) neurotransmission from MB-V3 neurons is necessary for LTM retrieval; and (v) RNAi-mediated down-regulation of oo18 RNA-binding protein (involved in local regulation of protein translation) in MB-V3 neurons impairs LTM. Our results suggest a model of long-term memory formation that includes a systems-level consolidation process, wherein an early, labile olfactory memory represented by neural activity in a sparse subset of MB neurons is converted into a stable LTM through protein synthesis in dendrites of MB-V3 neurons synapsed onto MB α lobes.
Keywords: CPEB; CREB; PUM; STAU; fragile X mental retardation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Ule J, Darnell RB. RNA binding proteins and the regulation of neuronal synaptic plasticity. Curr Opin Neurobiol. 2006;16(1):102–110. - PubMed
-
- Si K, Choi YB, White-Grindley E, Majumdar A, Kandel ER. Aplysia CPEB can form prion-like multimers in sensory neurons that contribute to long-term facilitation. Cell. 2010;140(3):421–435. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
