

Synthesis of customized petroleum-replica fuel molecules by targeted modification of free fatty acid pools in Escherichia coli
- PMID: 23610415
- PMCID: PMC3651483
- DOI: 10.1073/pnas.1215966110
Synthesis of customized petroleum-replica fuel molecules by targeted modification of free fatty acid pools in Escherichia coli
Abstract
Biofuels are the most immediate, practical solution for mitigating dependence on fossil hydrocarbons, but current biofuels (alcohols and biodiesels) require significant downstream processing and are not fully compatible with modern, mass-market internal combustion engines. Rather, the ideal biofuels are structurally and chemically identical to the fossil fuels they seek to replace (i.e., aliphatic n- and iso-alkanes and -alkenes of various chain lengths). Here we report on production of such petroleum-replica hydrocarbons in Escherichia coli. The activity of the fatty acid (FA) reductase complex from Photorhabdus luminescens was coupled with aldehyde decarbonylase from Nostoc punctiforme to use free FAs as substrates for alkane biosynthesis. This combination of genes enabled rational alterations to hydrocarbon chain length (Cn) and the production of branched alkanes through upstream genetic and exogenous manipulations of the FA pool. Genetic components for targeted manipulation of the FA pool included expression of a thioesterase from Cinnamomum camphora (camphor) to alter alkane Cn and expression of the branched-chain α-keto acid dehydrogenase complex and β-keto acyl-acyl carrier protein synthase III from Bacillus subtilis to synthesize branched (iso-) alkanes. Rather than simply reconstituting existing metabolic routes to alkane production found in nature, these results demonstrate the ability to design and implement artificial molecular pathways for the production of renewable, industrially relevant fuel molecules.
Keywords: branched fatty acid biosynthesis; lux genes; metabolic engineering; synthetic biology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
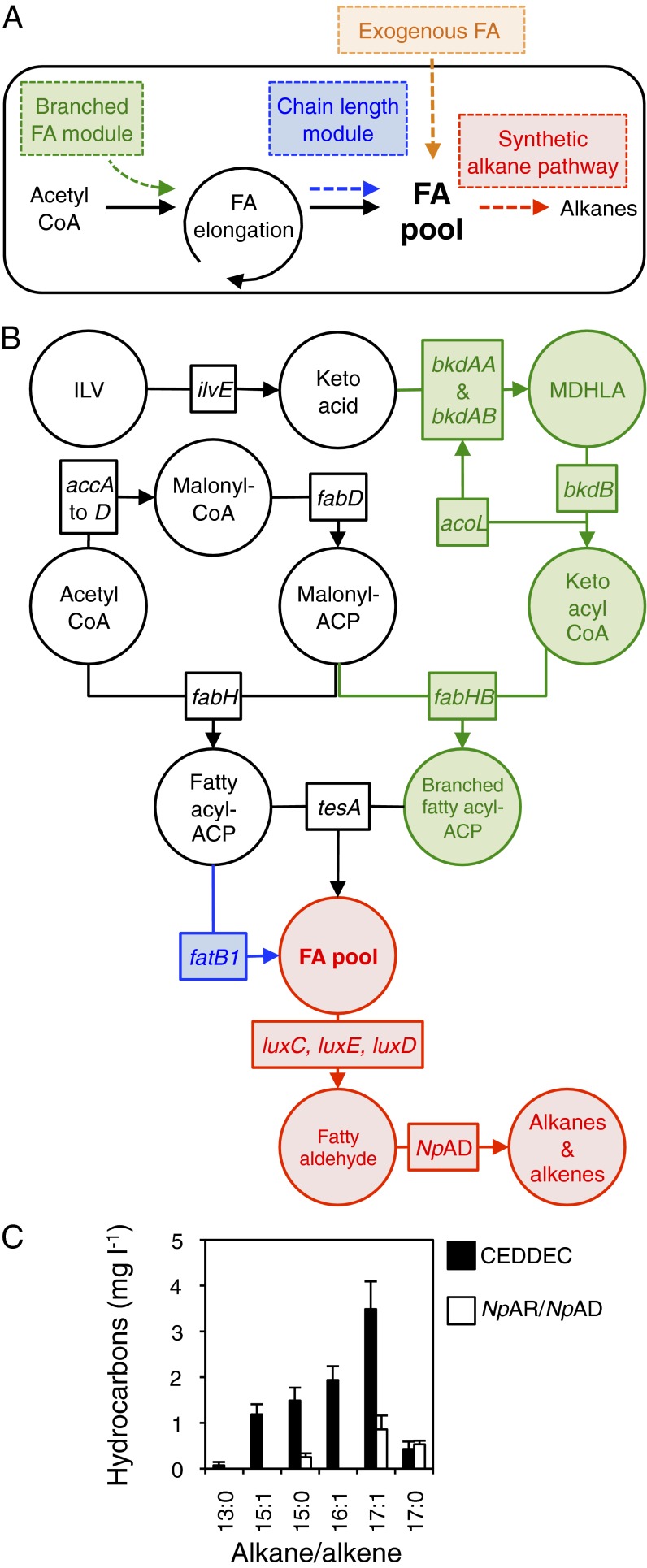
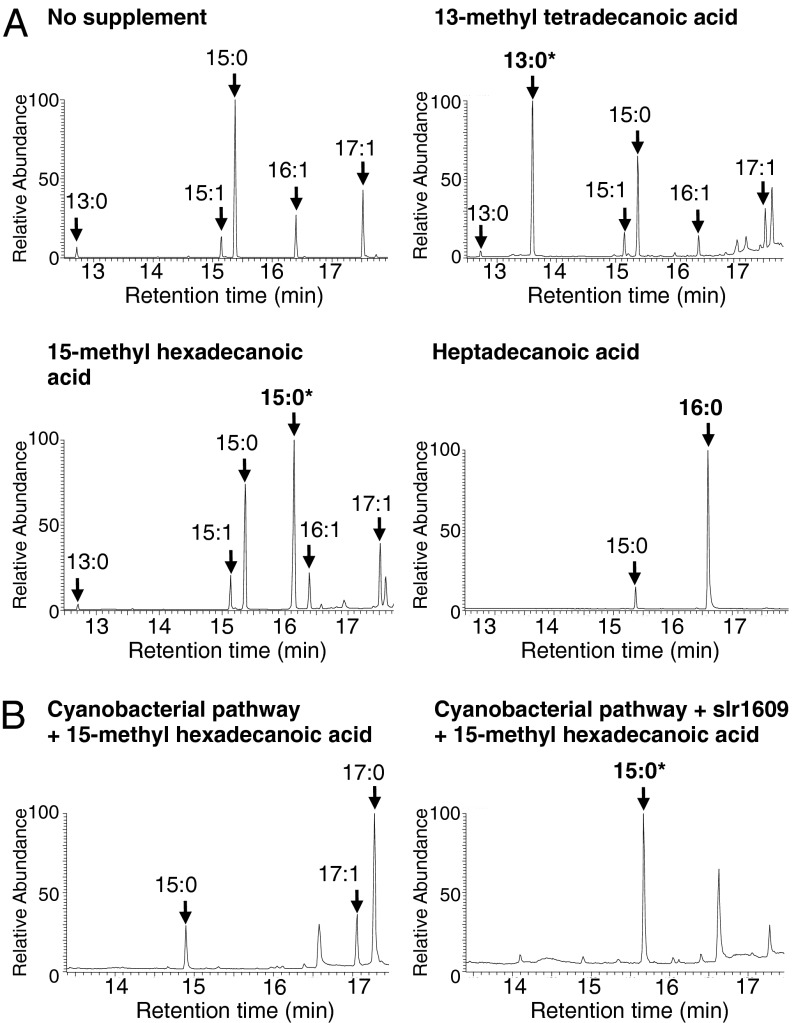
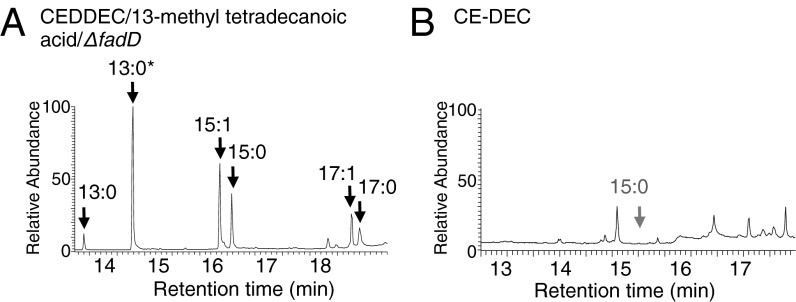
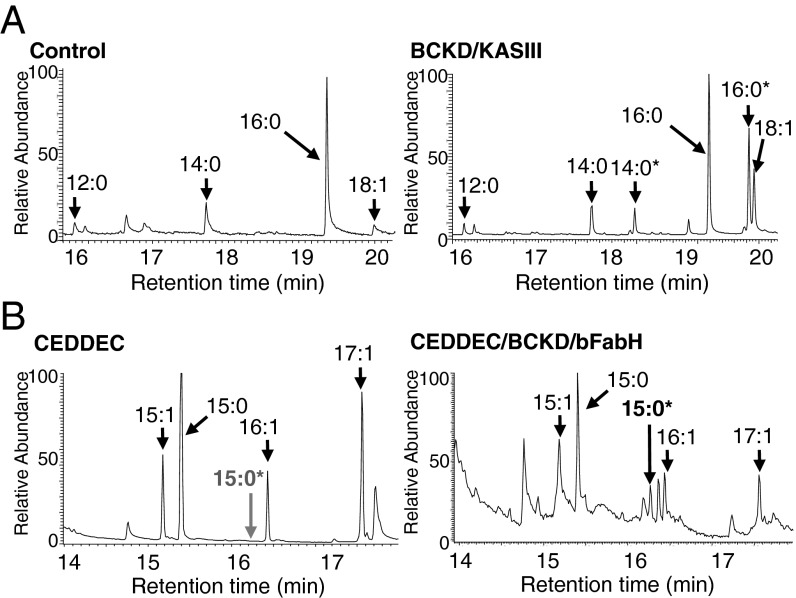
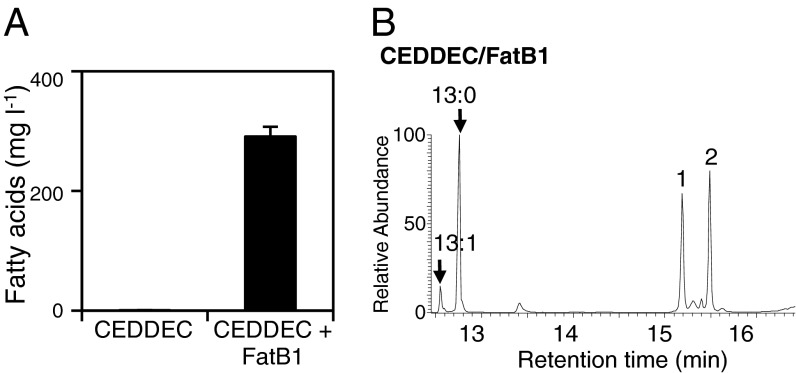
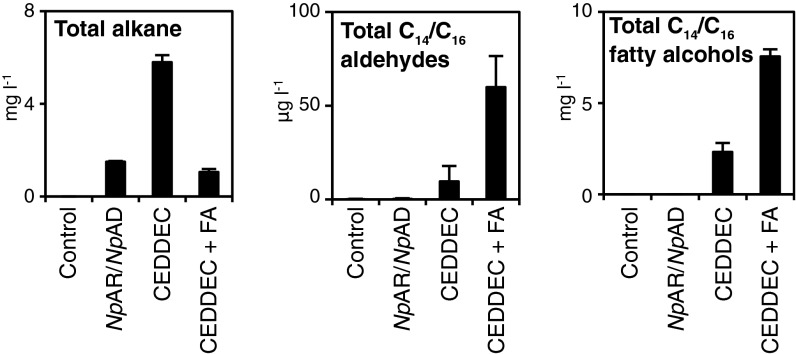
Comment in
-
Industrial microbiology: Designer biofuels?Nat Rev Microbiol. 2013 Jun;11(6):360. doi: 10.1038/nrmicro3044. Nat Rev Microbiol. 2013. PMID: 23676927 No abstract available.
References
-
- International Energy Agency . Key World Statistics. Paris, France: 2011.
-
- International Energy Agency . World Energy Outlook. Paris, France: 2008.
-
- European Directive (2009) Council Directive (EC) 2009/28/EC of 23 April 2009 on the promotion of the use of energy from renewable sources and amending and subsequently repealing Directives 2001/77/EC and 2003/30/EC.
-
- National Renewable Energy Laboratory . Biodiesel Handling and User Guide. 4th Ed. US Department of Commerce: Oak Ridge, TN; 2009.
-
- Yuksel F, Yuksel B. The use of ethanol-gasoline blend as a fuel in an SI engine. Renew Energy. 2004;29:1181–1191.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
