

Beyond TGFβ: roles of other TGFβ superfamily members in cancer
- PMID: 23612460
- PMCID: PMC7608560
- DOI: 10.1038/nrc3500
Beyond TGFβ: roles of other TGFβ superfamily members in cancer
Abstract
Much of the focus on the transforming growth factor-β (TGFβ) superfamily in cancer has revolved around the TGFβ ligands themselves. However, it is now becoming apparent that deregulated signalling by many of the other superfamily members also has crucial roles in both the development of tumours and metastasis. Furthermore, these signalling pathways are emerging as plausible therapeutic targets. Their roles in tumorigenesis frequently reflect their function in embryonic development or in adult tissue homeostasis, and their influence extends beyond the tumours themselves, to the tumour microenvironment and more widely to complications of cancer such as cachexia and bone loss.
Conflict of interest statement
Competing interests statement
The authors declare no competing financial interests.
Figures
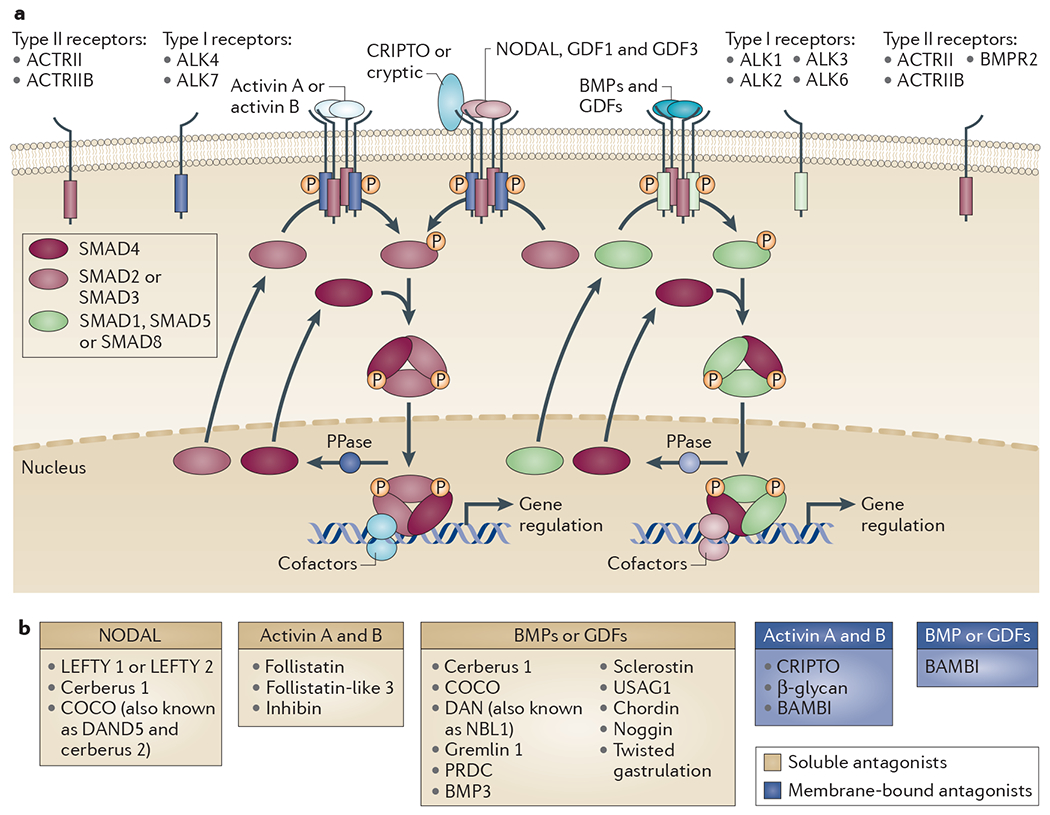
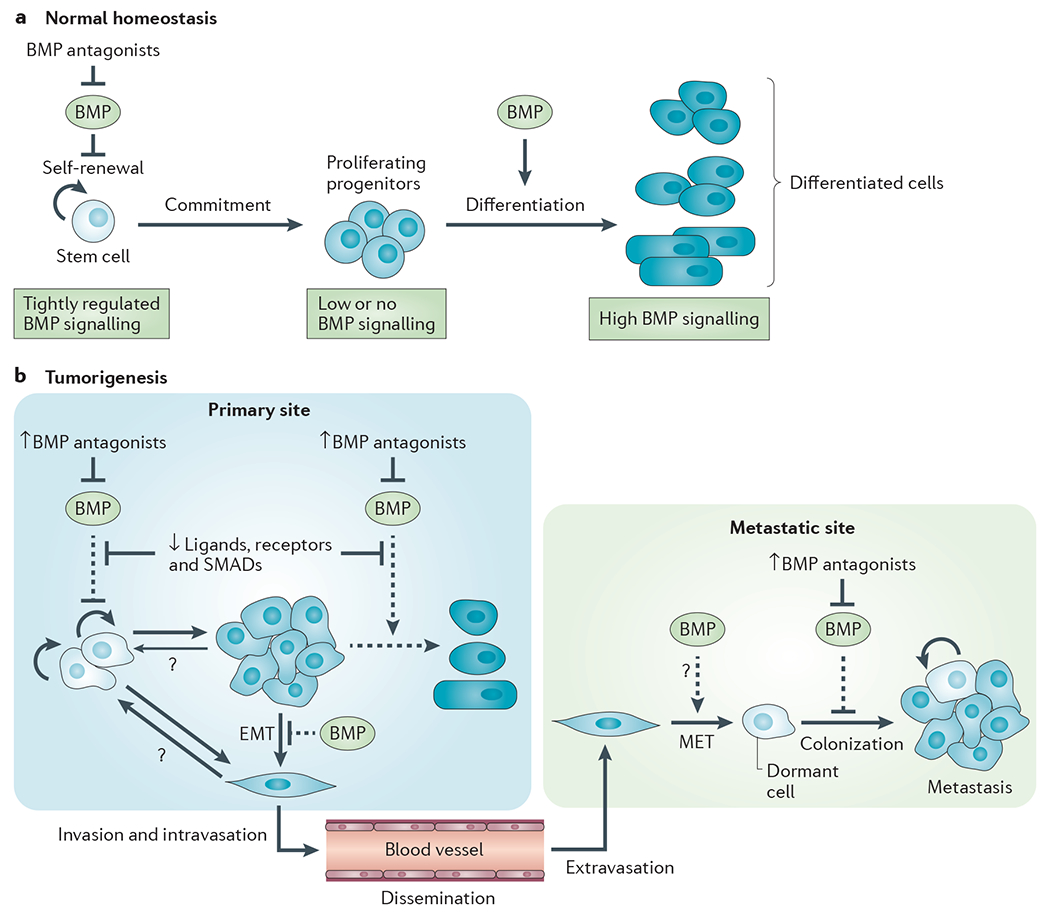
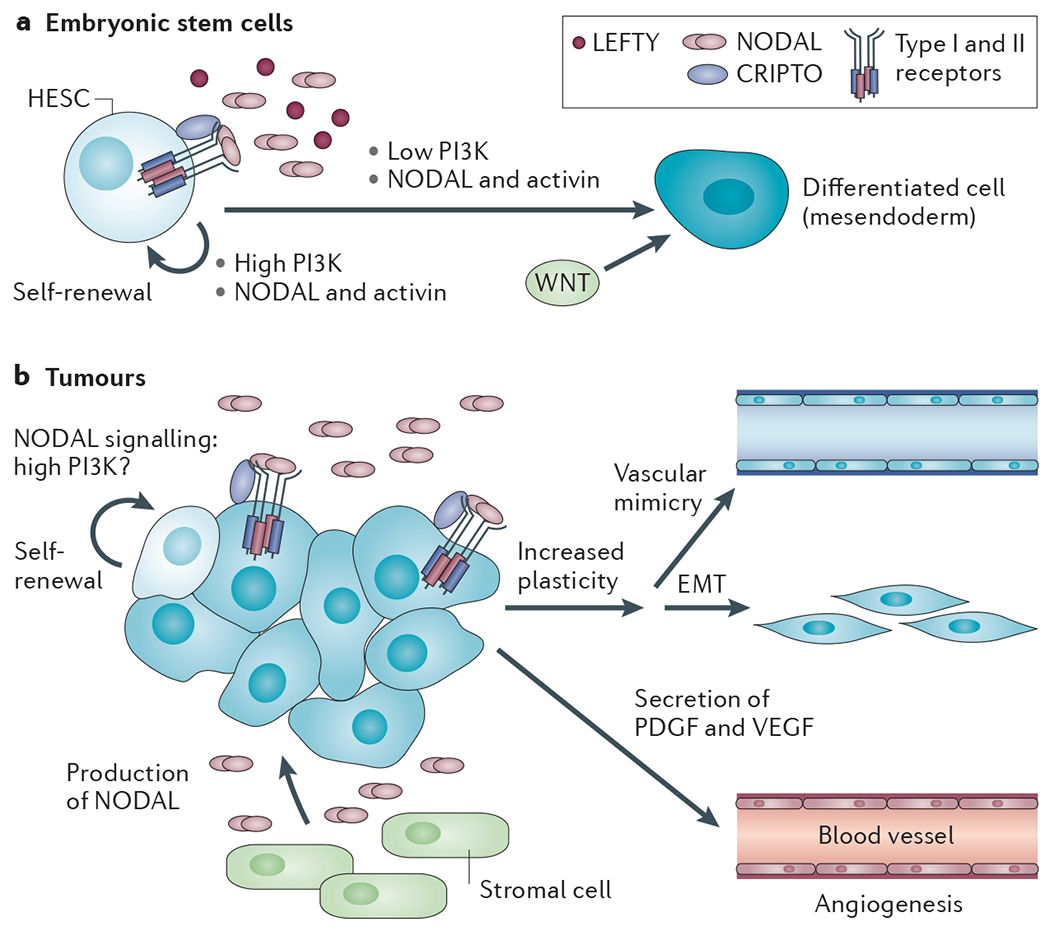
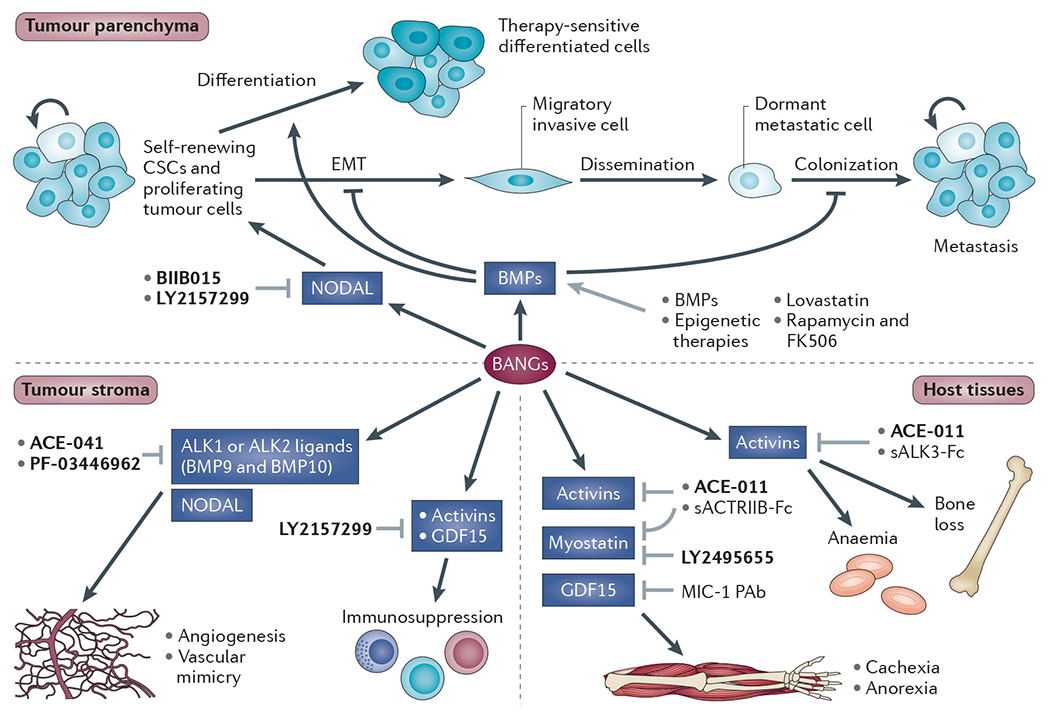
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
