

Stabilization of integrin-linked kinase by the Hsp90-CHIP axis impacts cellular force generation, migration and the fibrotic response
- PMID: 23612611
- PMCID: PMC3655474
- DOI: 10.1038/emboj.2013.90
Stabilization of integrin-linked kinase by the Hsp90-CHIP axis impacts cellular force generation, migration and the fibrotic response
Abstract
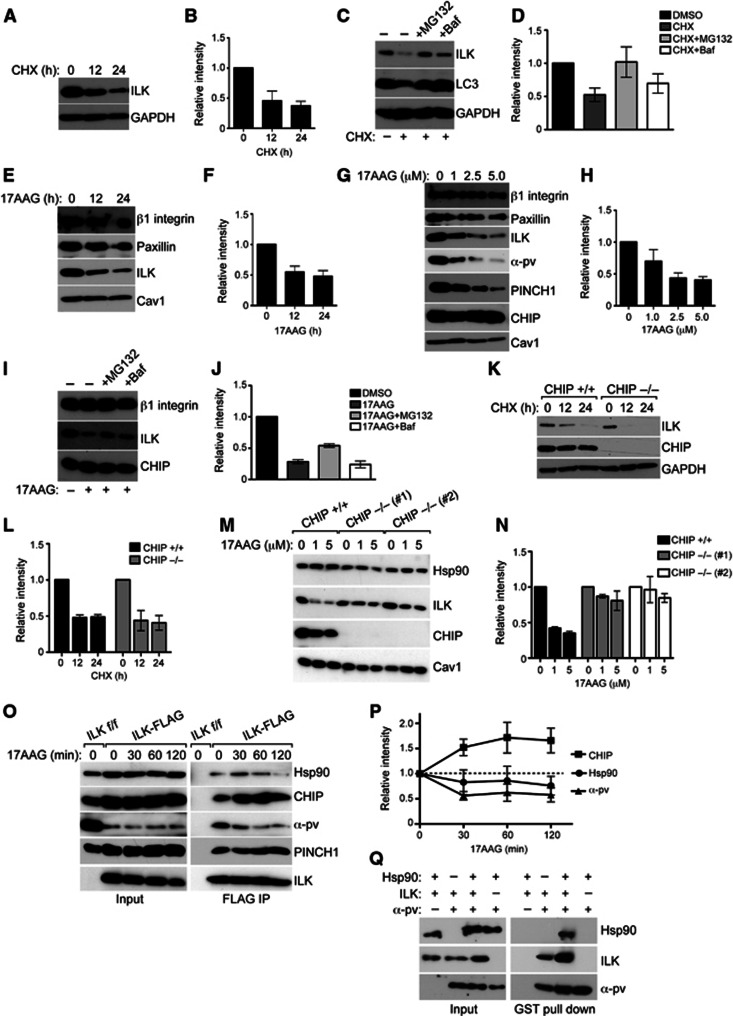
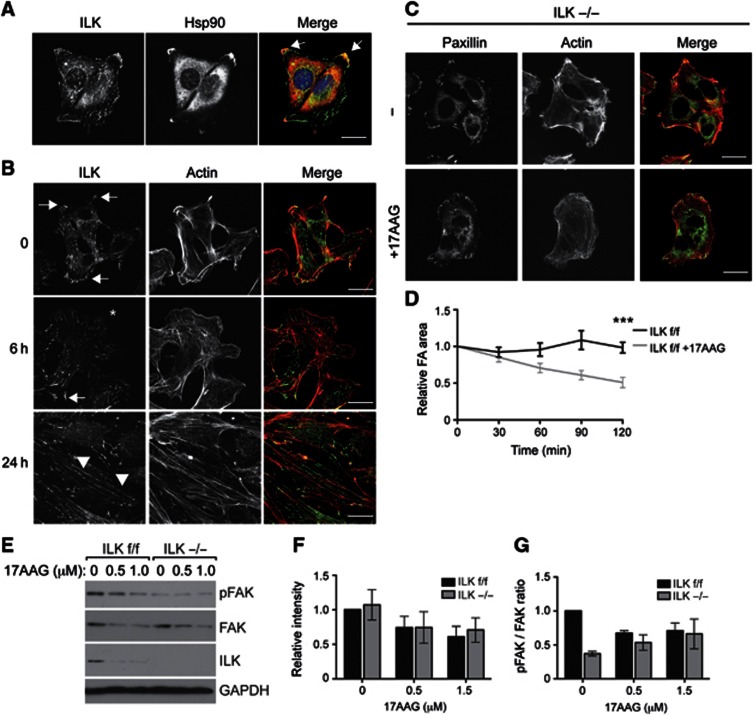
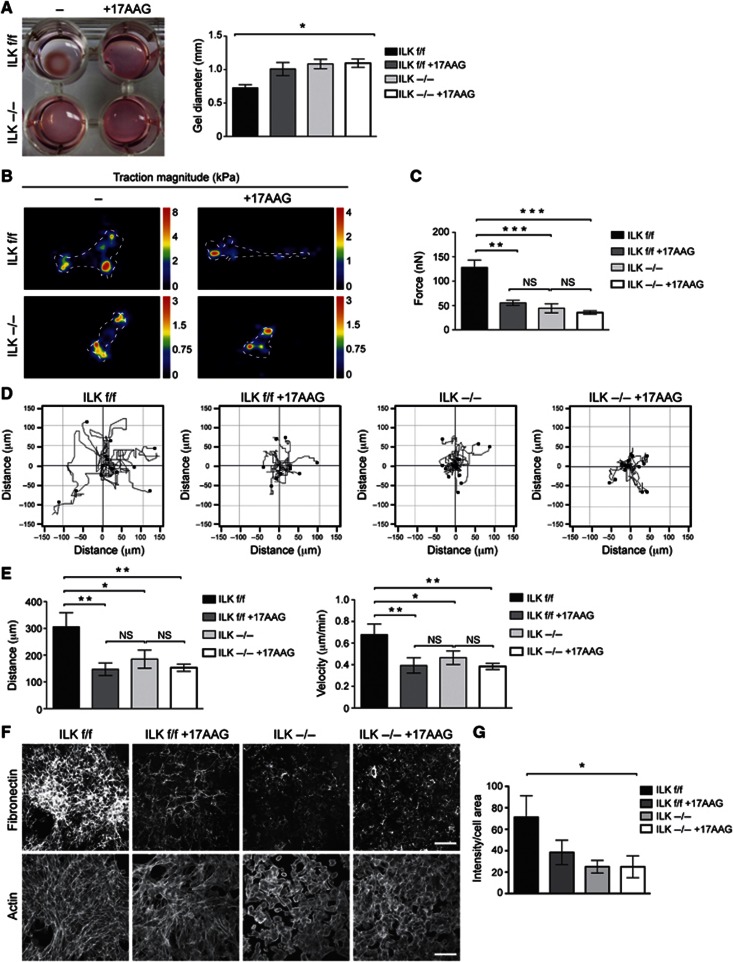
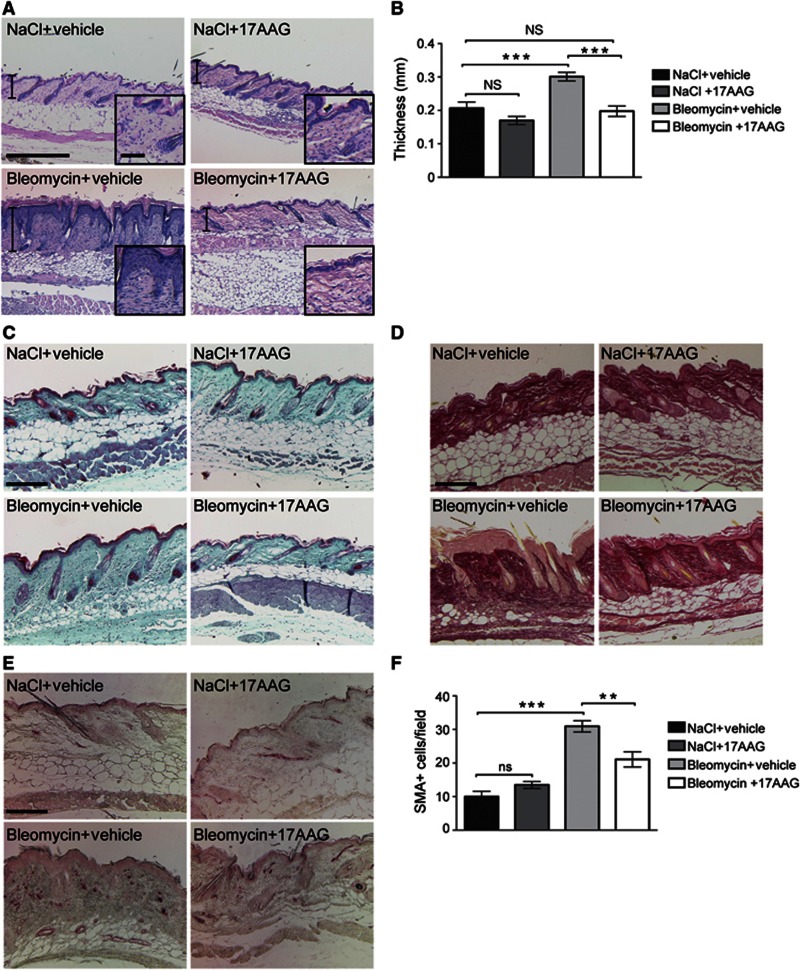
Integrin-linked kinase (ILK) is an adaptor protein required to establish and maintain the connection between integrins and the actin cytoskeleton. This linkage is essential for generating force between the extracellular matrix (ECM) and the cell during migration and matrix remodelling. The mechanisms by which ILK stability and turnover are regulated are unknown. Here we report that the E3 ligase CHIP-heat shock protein 90 (Hsp90) axis regulates ILK turnover in fibroblasts. The chaperone Hsp90 stabilizes ILK and facilitates the interaction of ILK with α-parvin. When Hsp90 activity is blocked, ILK is ubiquitinated by CHIP and degraded by the proteasome, resulting in impaired fibroblast migration and a dramatic reduction in the fibrotic response to bleomycin in mice. Together, our results uncover how Hsp90 regulates ILK stability and identify a potential therapeutic strategy to alleviate fibrotic diseases.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
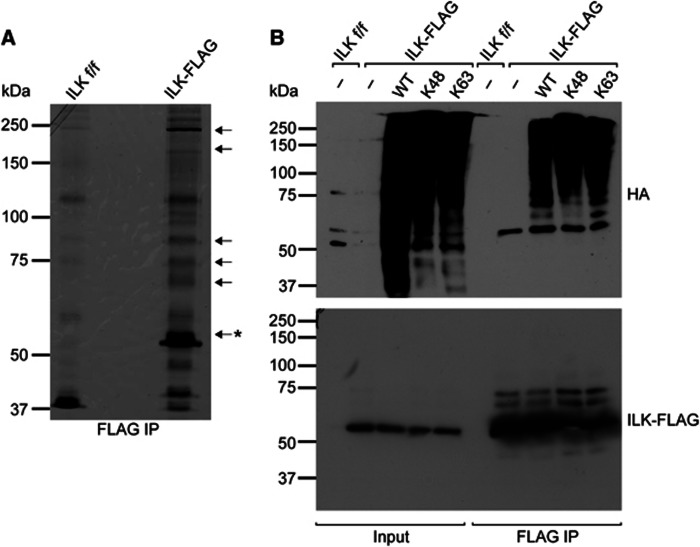
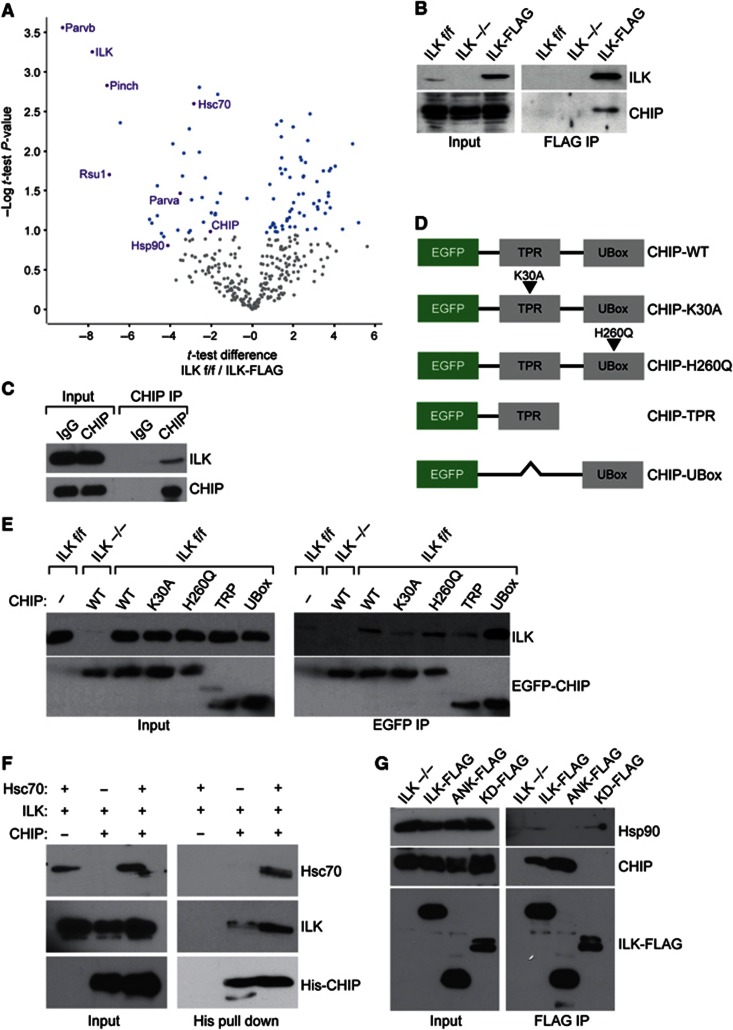
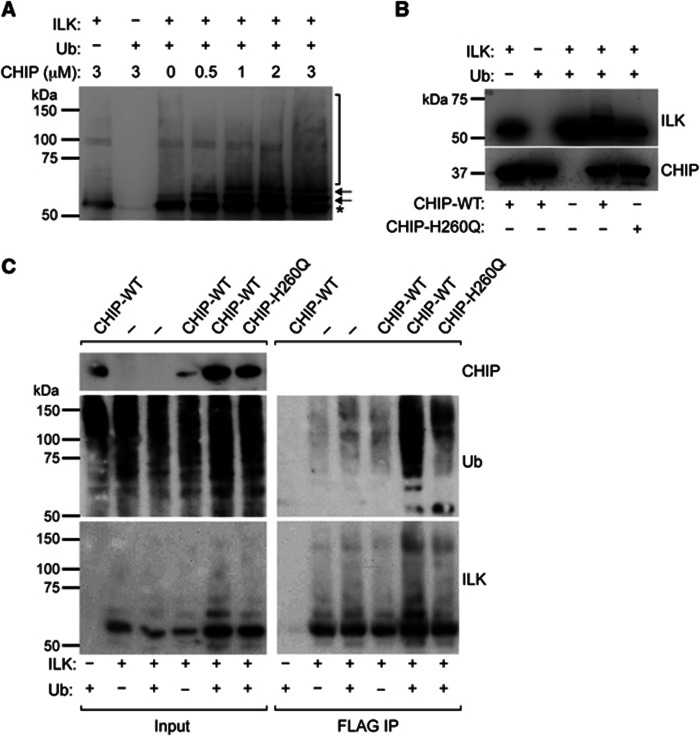
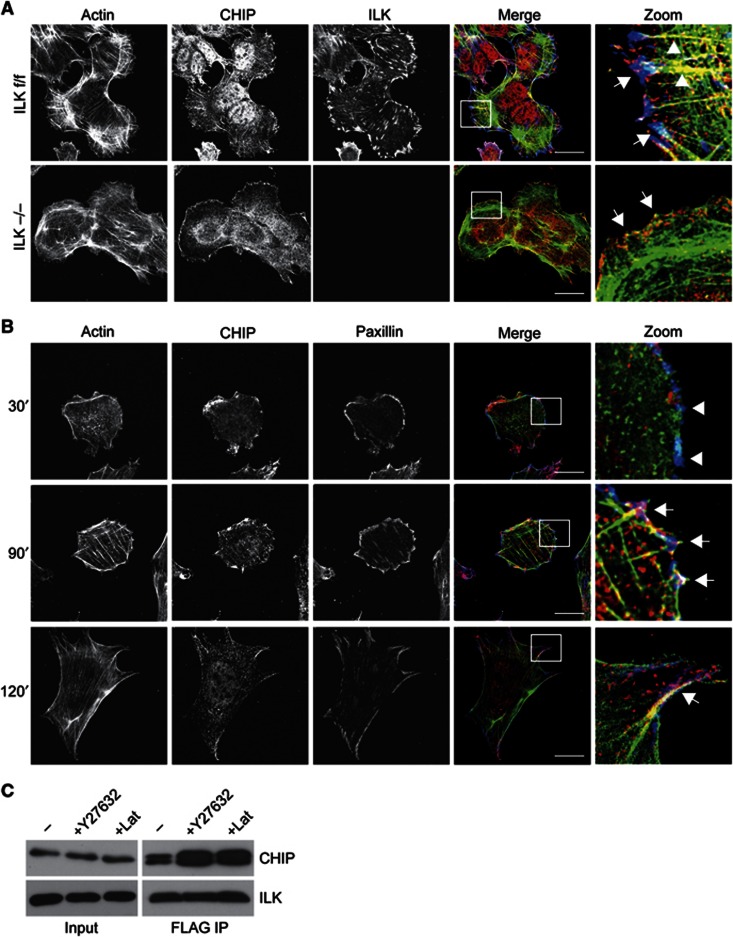
Comment in
-
The weakest link: a new paradigm for stabilizing the integrin-actin connection.Cell Cycle. 2013 Sep 15;12(18):2929-30. doi: 10.4161/cc.26213. Epub 2013 Aug 23. Cell Cycle. 2013. PMID: 23974089 Free PMC article. No abstract available.
References
-
- Aoyagi Y, Fujita N, Tsuruo T (2005) Stabilization of integrin-linked kinase by binding to Hsp90. Biochem Biophys Res Commun 331: 1061–1068 - PubMed
-
- Arndt V, Dick N, Tawo R, Dreiseidler M, Wenzel D, Hesse M, Furst DO, Saftig P, Saint R, Fleischmann BK, Hoch M, Höhfeld J (2010) Chaperone-assisted selective autophagy is essential for muscle maintenance. Curr Biol 20: 143–148 - PubMed
-
- Azimifar SB, Böttcher RT, Zanivan S, Grashoff C, Kruger M, Legate KR, Mann M, Fässler R (2012) Induction of membrane circular dorsal ruffles requires co-signalling of integrin-ILK-complex and EGF receptor. J Cell Sci 125: 435–448 - PubMed
-
- Blumbach K, Zweers MC, Brunner G, Peters AS, Schmitz M, Schulz JN, Schild A, Denton CP, Sakai T, Fässler R, Krieg T, Eckes B (2010) Defective granulation tissue formation in mice with specific ablation of integrin-linked kinase in fibroblasts—role of TGFbeta1 levels and RhoA activity. J Cell Sci 123: 3872–3883 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
