

Pheromone-induced morphogenesis improves osmoadaptation capacity by activating the HOG MAPK pathway
- PMID: 23612707
- PMCID: PMC3701258
- DOI: 10.1126/scisignal.2003312
Pheromone-induced morphogenesis improves osmoadaptation capacity by activating the HOG MAPK pathway
Abstract
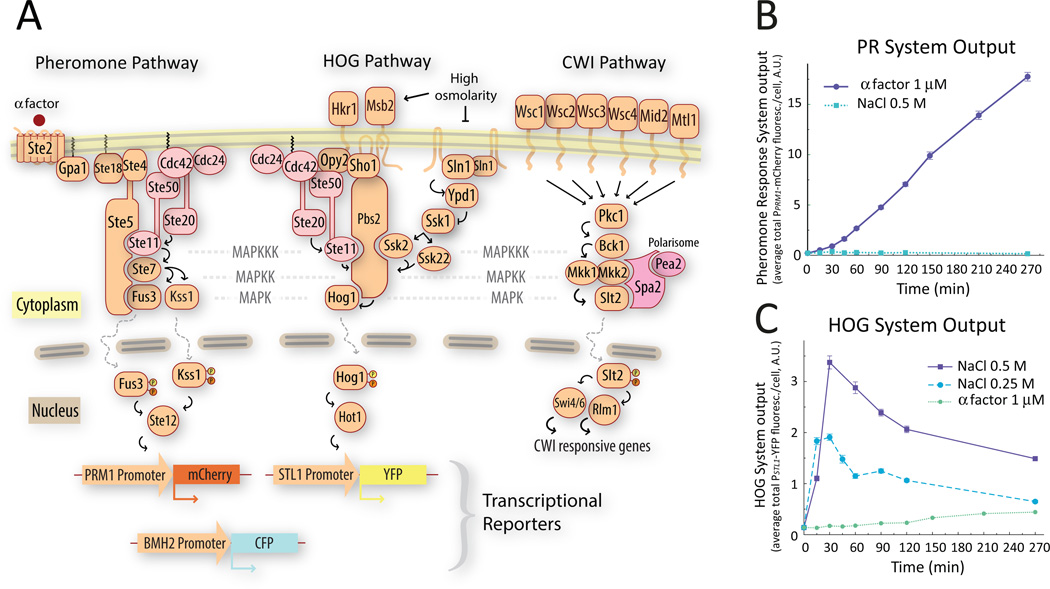
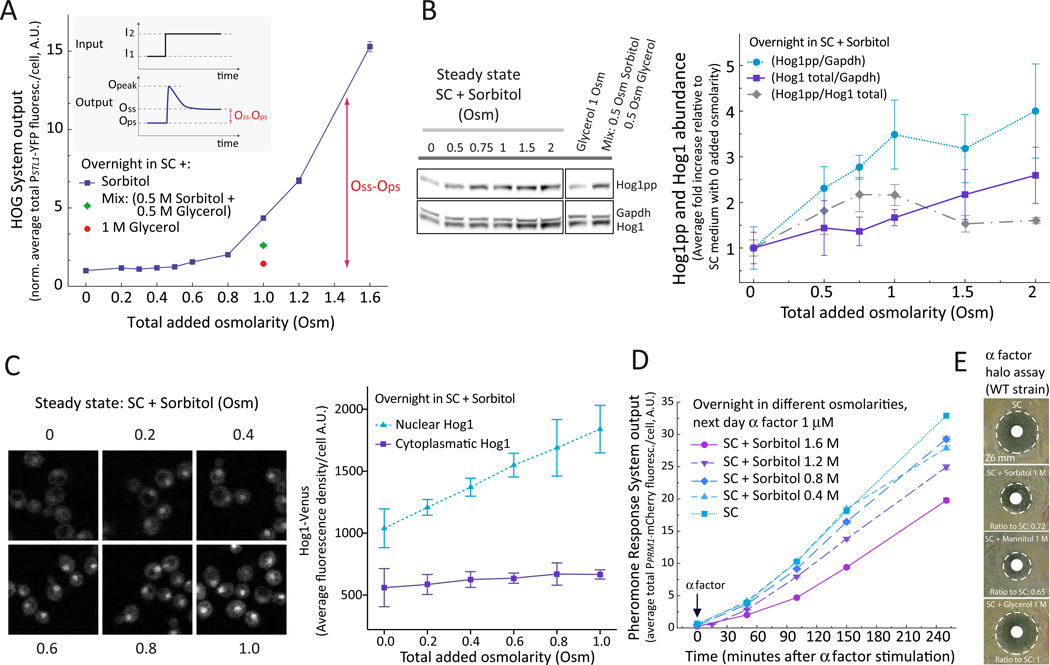
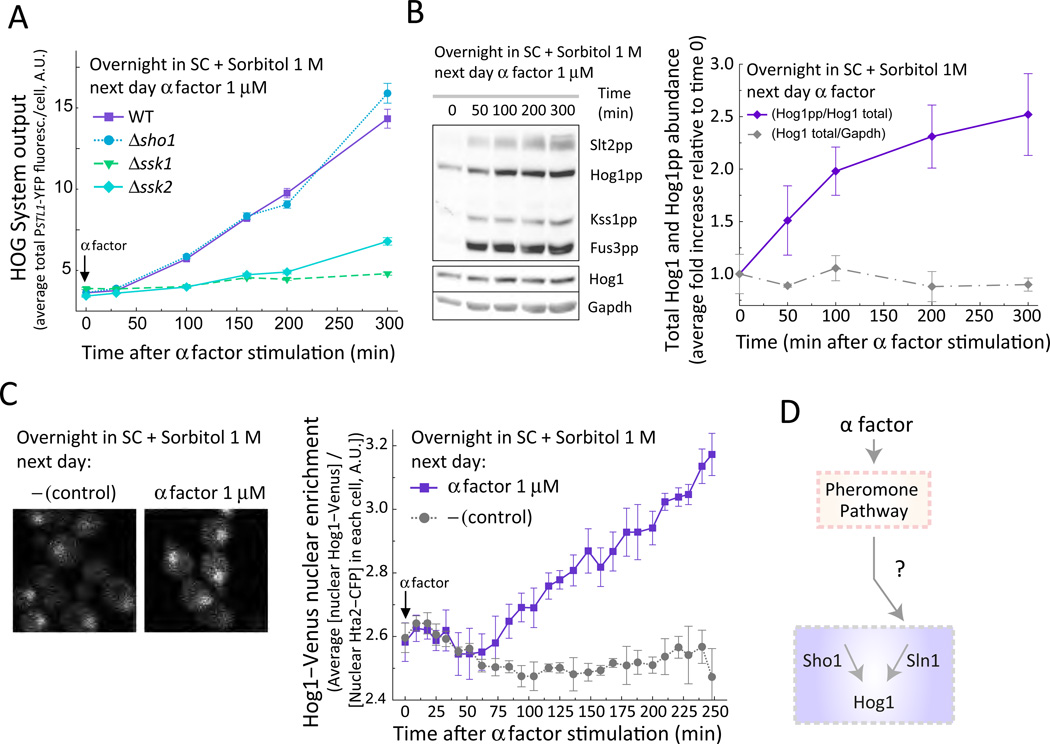
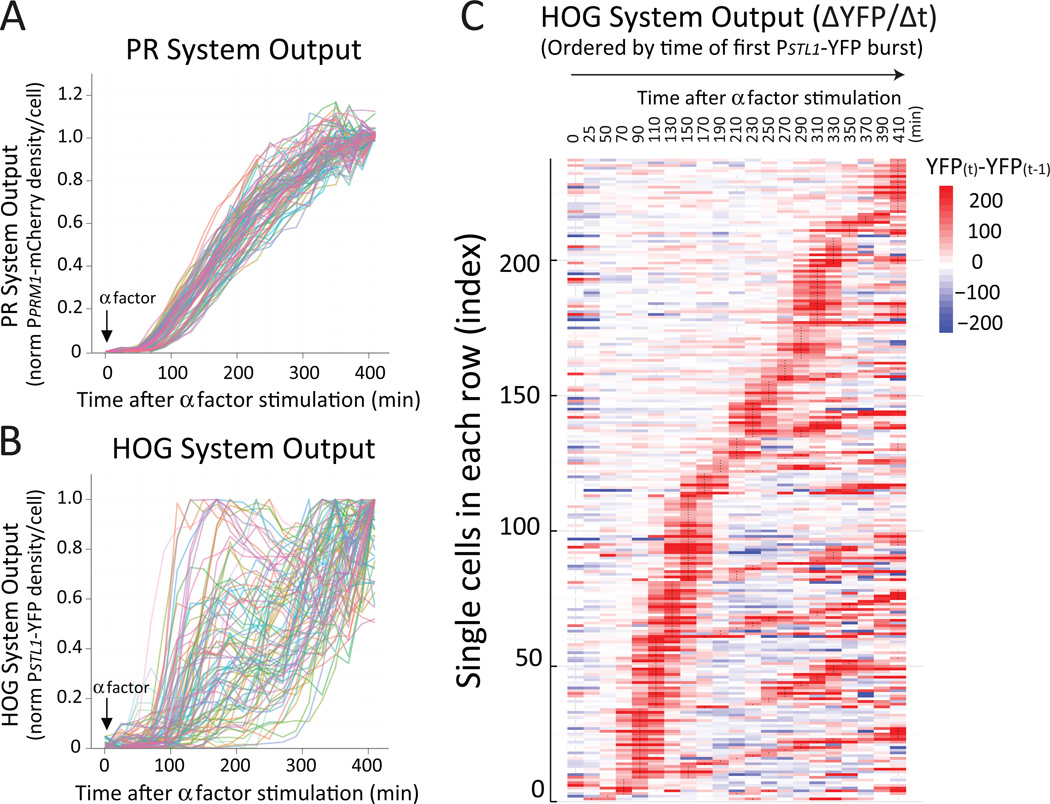
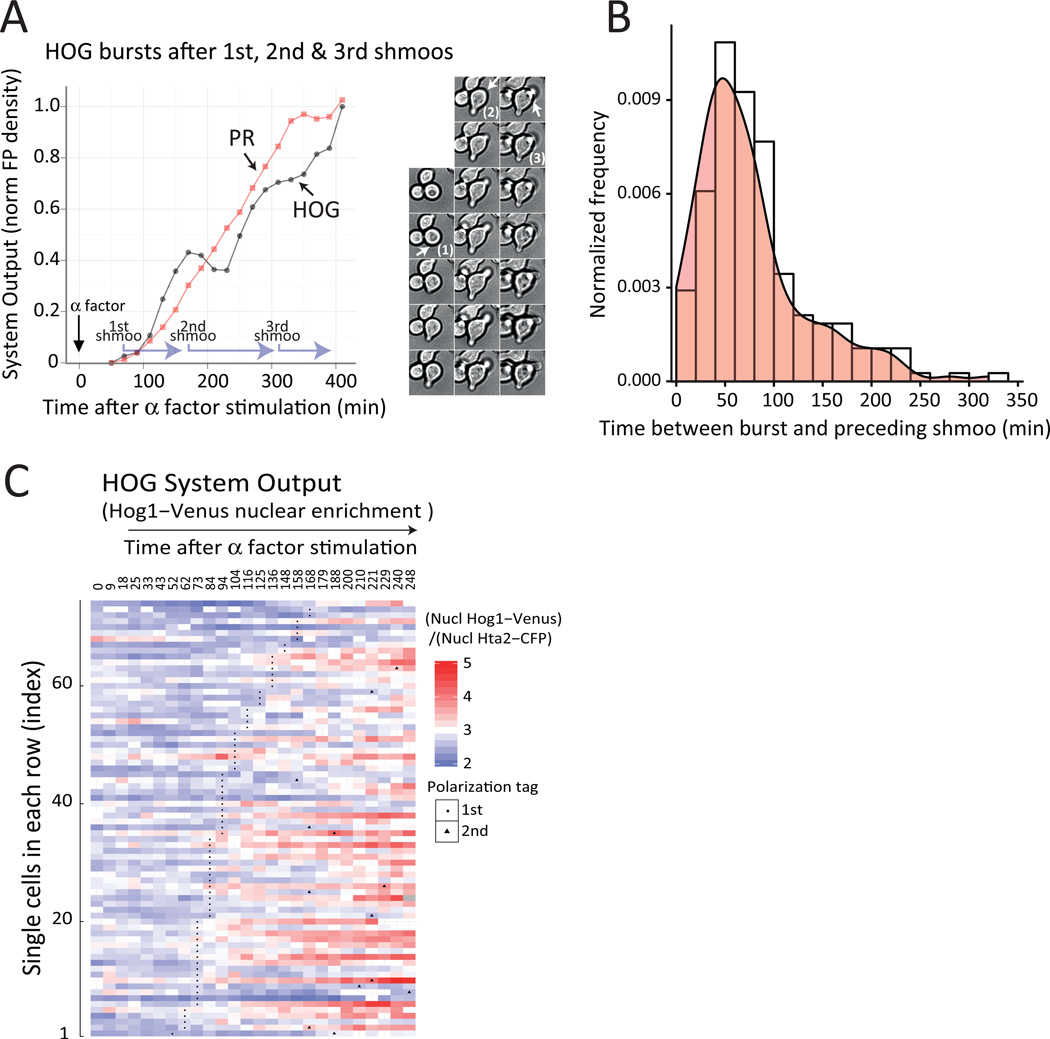
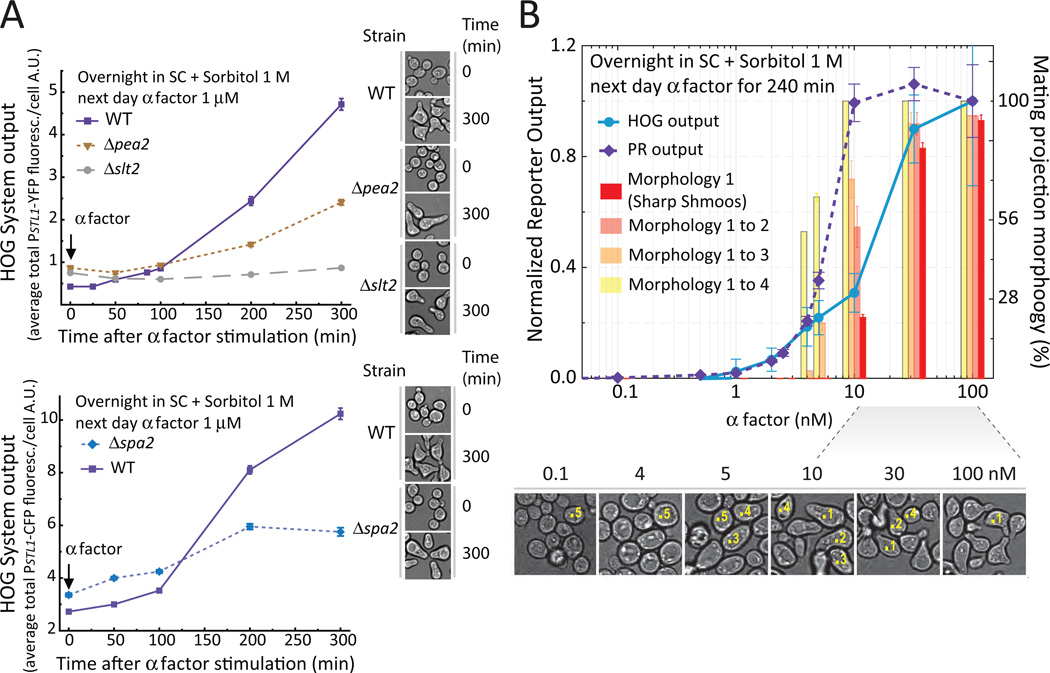
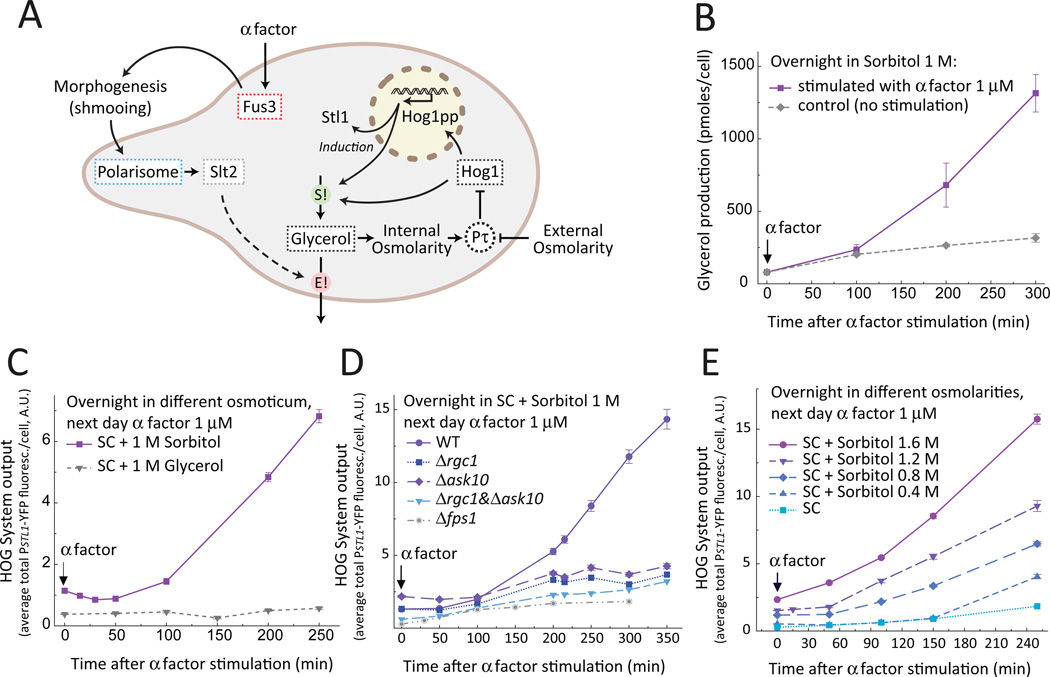
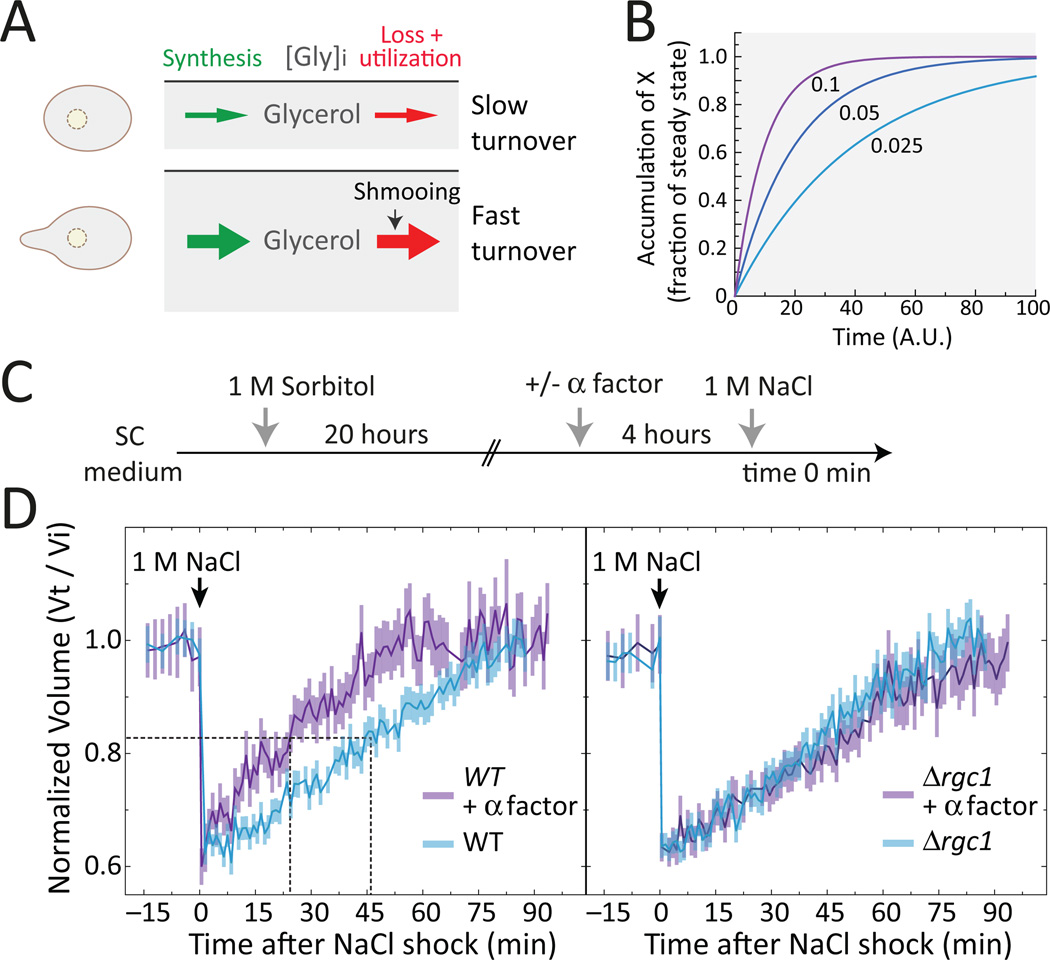
Environmental and internal conditions expose cells to a multiplicity of stimuli whose consequences are difficult to predict. We investigate the response to mating pheromone of yeast cells adapted to high osmolarity. Events downstream of pheromone binding involve two mitogen-activated protein kinase (MAPK) cascades: the pheromone response (PR) and the cell wall integrity (CWI) response. Although the PR MAPK pathway shares components with a third MAPK pathway, the high osmolarity (HOG) response, each one is normally only activated by its cognate stimulus, a phenomenon called insulation. We found that in cells adapted to high osmolarity, PR activated the HOG pathway in a pheromone- and osmolarity-dependent manner. Activation of HOG by the PR was not due to loss of insulation, but rather a response to a reduction in internal osmolarity, which resulted from an increase in glycerol release caused by the PR. By analyzing single-cell time courses, we found that stimulation of HOG occurred in discrete bursts that coincided with the "shmooing" morphogenetic process. Activation required the polarisome, the CWI MAPK Slt2, and the aquaglyceroporin Fps1. HOG activation resulted in high glycerol turnover, which improved adaptability to rapid changes in osmolarity. Our work shows how a differentiation signal can recruit a second, unrelated sensory pathway to fine-tune yeast response in a complex environment.
Figures
References
-
- Dohlman HG, Thorner JW. Regulation of G protein-initiated signal transduction in yeast: Paradigms and Principles. Annu.Rev.Biochem. 2001;70:703. - PubMed
-
- Madhani HD, Fink GR. The control of filamentous differentiation and virulence in fungi. Trends Cell Biol. 1998;8:348. - PubMed
-
- Roberts CJ, Nelson B, Marton MJ, Stoughton R, Meyer MR, Bennett HA, He YD, Dai H, Walker WL, Hughes TR, Tyers M, Boone C, Friend SH. Signaling and circuitry of multiple MAPK pathways revealed by a matrix of global gene expression profiles. Science. 2000;287:873. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
