

Kinetic mechanism of human histidine triad nucleotide binding protein 1
- PMID: 23614568
- PMCID: PMC3835729
- DOI: 10.1021/bi301616c
Kinetic mechanism of human histidine triad nucleotide binding protein 1
Abstract
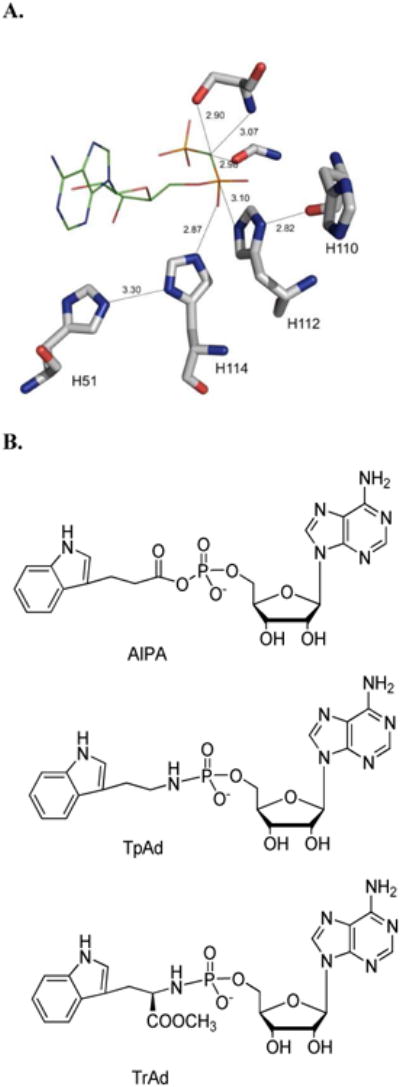
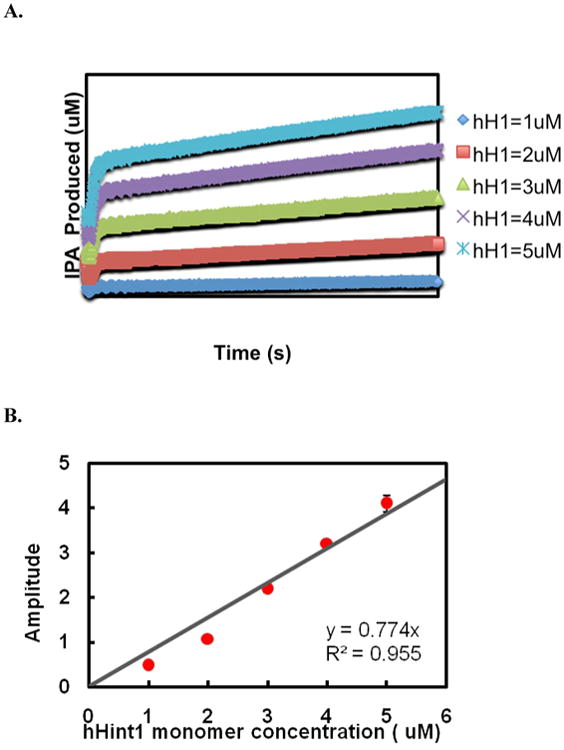
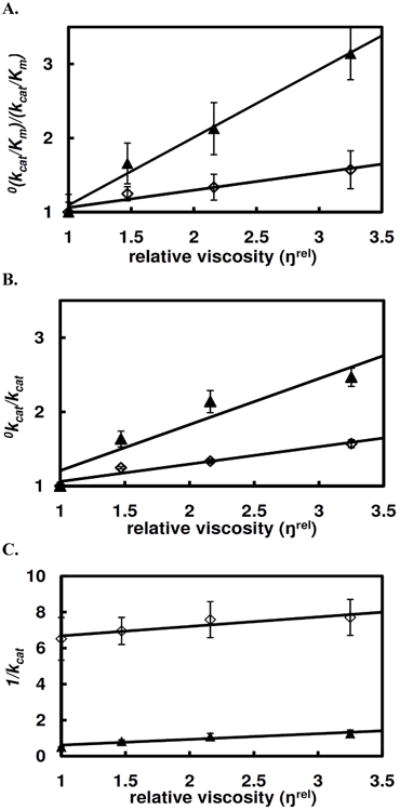
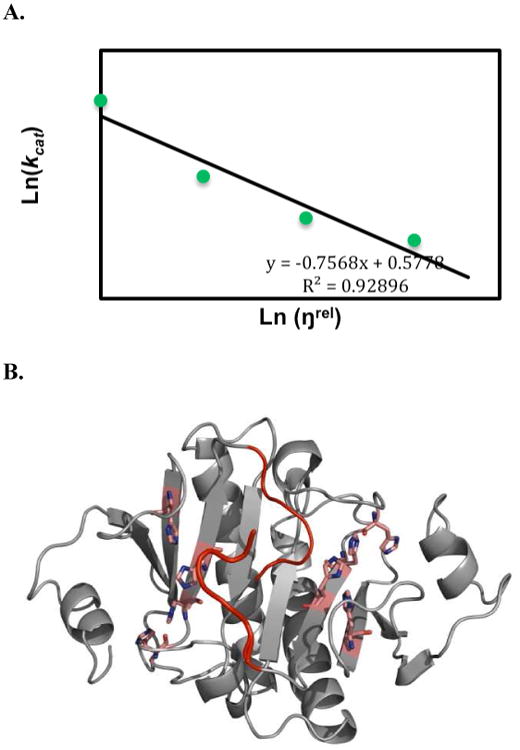
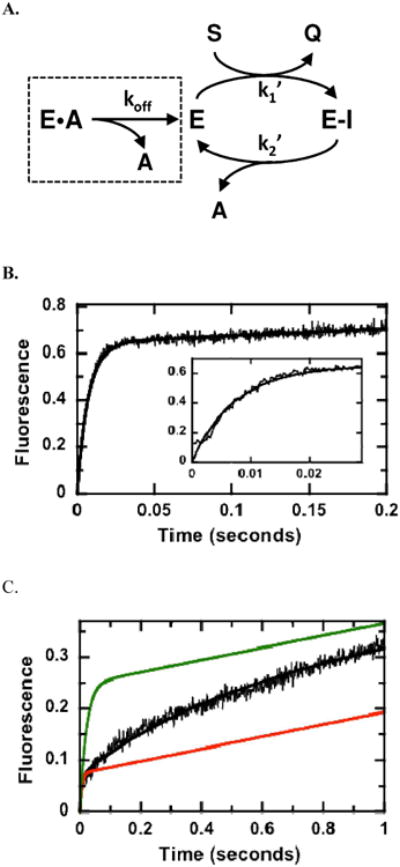
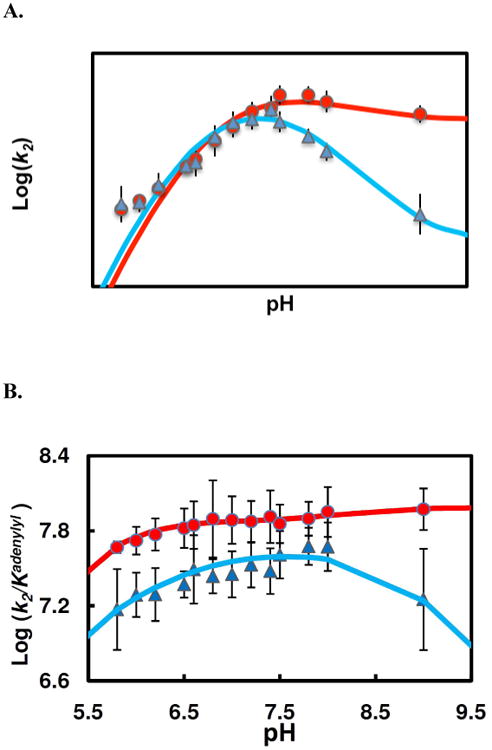
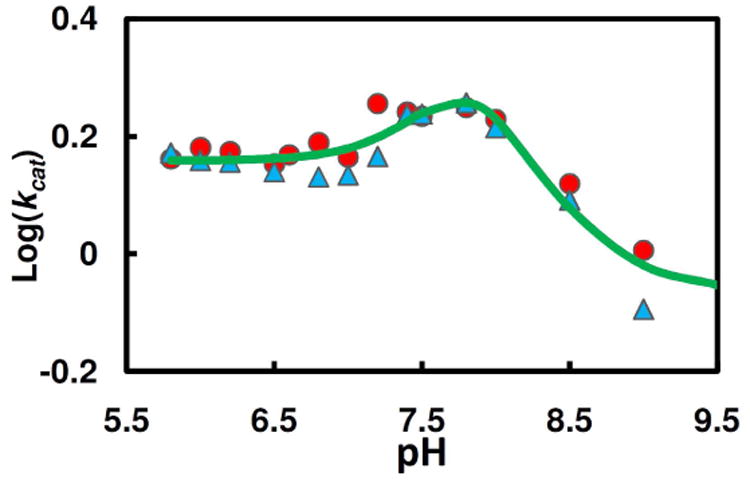
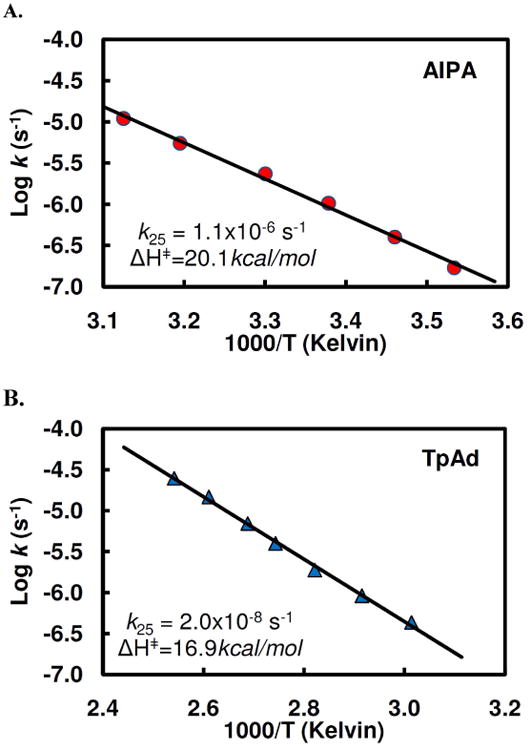
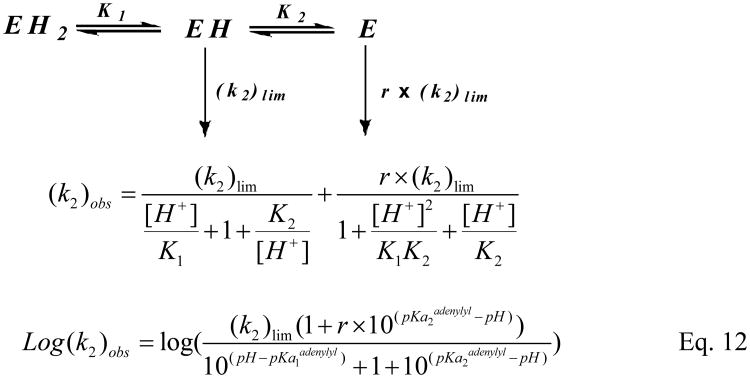
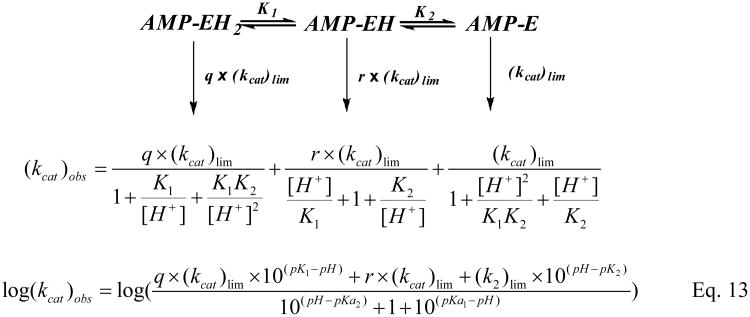
Human histidine triad nucleotide binding protein 1 (hHint1) is a member of a ubiquitous and ancient branch of the histidine triad protein superfamily. hHint1 is a homodimeric protein that catalyzes the hydrolysis of model substrates, phosphoramidate and acyl adenylate, with a high efficiency. Recently, catalytically inactive hHint1 has been identified as the cause of inherited peripheral neuropathy [Zimon, M., et al. (2012) Nat. Genet. 44, 1080-1083]. We have conducted the first detailed kinetic mechanistic studies of hHint1 and have found that the reaction mechanism is consistent with a double-displacement mechanism, in which the active site nucleophile His112 is first adenylylated by the substrate, followed by hydrolysis of the AMP-enzyme intermediate. A transient burst phase followed by a linear phase from the stopped-flow fluorescence assay indicated that enzyme adenylylation was faster than the subsequent intermediate hydrolysis and product release. Solvent viscosity experiments suggested that both chemical transformation and diffusion-sensitive events (product release or protein conformational change) limit the overall turnover. The catalytic trapping experiments and data simulation indicated that the true koff rate of the final product AMP is unlikely to control the overall kcat. Therefore, a protein conformational change associated with product release is likely rate-limiting. In addition, the rate of Hint1 adenylylation was found to be dependent on two residues with pKa values of 6.5 and 8, with the former pKa agreeing well with the nuclear magnetic resonance titration results for the pKa of the active site nucleophile His112. In comparison to the uncatalyzed rates, hHint1 was shown to enhance acyl-AMP and AMP phosphoramidate hydrolysis by 10(6)-10(8)-fold. Taken together, our analysis indicates that hHint1 catalyzes the hydrolysis of phosphoramidate and acyl adenylate with high efficiency, through a mechanism that relies on rapid adenylylation of the active residue, His112, while being partially rate-limited by intermediate hydrolysis and product release associated with a conformational change. Given the high degree of sequence homology of Hint proteins across all kingdoms of life, it is likely that their kinetic and catalytic mechanisms will be similar to those elucidated for hHint1.
Figures
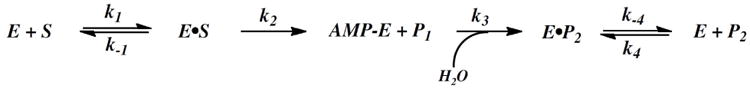

References
-
- Zimon M, Baets J, Almeida-Souza L, De Vriendt E, Nikodinovic J, Parman Y, Battaloglu E, Matur Z, Guergueltcheva V, Tournev I, Auer-Grumbach M, De Rijk P, Petersen BS, Muller T, Fransen E, Van Damme P, Loscher WN, Barisic N, Mitrovic Z, Previtali SC, Topaloglu H, Bernert G, Beleza-Meireles A, Todorovic S, Savic-Pavicevic D, Ishpekova B, Lechner S, Peeters K, Ooms T, Hahn AF, Zuchner S, Timmerman V, Van Dijck P, Rasic VM, Janecke AR, De Jonghe P, Jordanova A. Loss-of-function mutations in HINT1 cause axonal neuropathy with neuromyotonia. Nat Genet. 2012;44:1080–1083. - PubMed
-
- Mozier NM, Walsh MP, Pearson JD. Characterization of a novel zinc binding site of protein kinase C inhibitor-1. FEBS Lett. 1991;279:14–18. - PubMed
-
- Rodriguez-Munoz M, de la Torre-Madrid E, Sanchez-Blazquez P, Wang JB, Garzon J. NMDAR-nNOS generated zinc recruits PKCgamma to the HINT1-RGS17 complex bound to the C terminus of Mu-opioid receptors. Cell Signal. 2008;20:1855–1864. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
