

Activation of ADF/cofilin by phosphorylation-regulated Slingshot phosphatase is required for the meiotic spindle assembly in Xenopus laevis oocytes
- PMID: 23615437
- PMCID: PMC3681698
- DOI: 10.1091/mbc.E12-12-0851
Activation of ADF/cofilin by phosphorylation-regulated Slingshot phosphatase is required for the meiotic spindle assembly in Xenopus laevis oocytes
Abstract
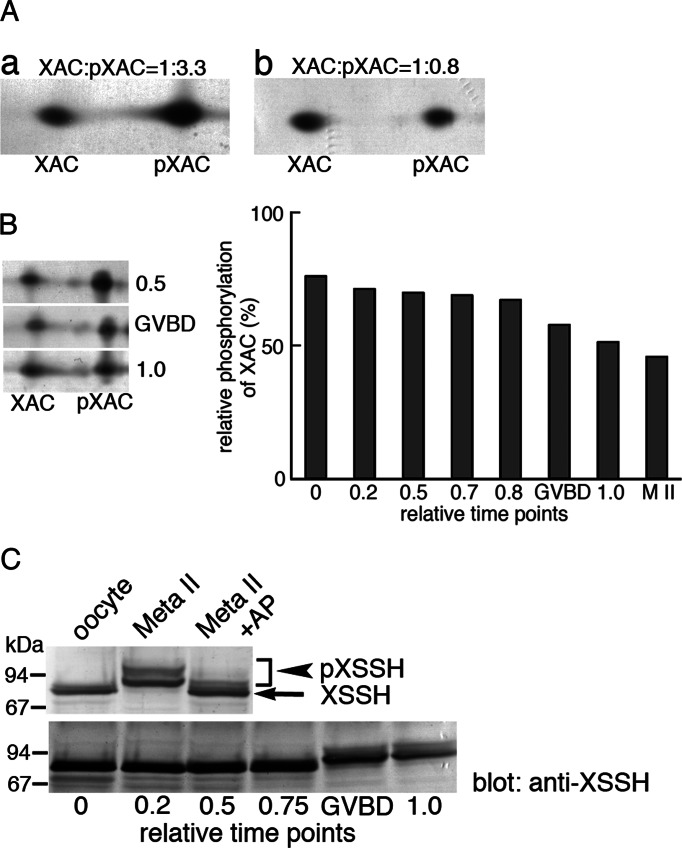
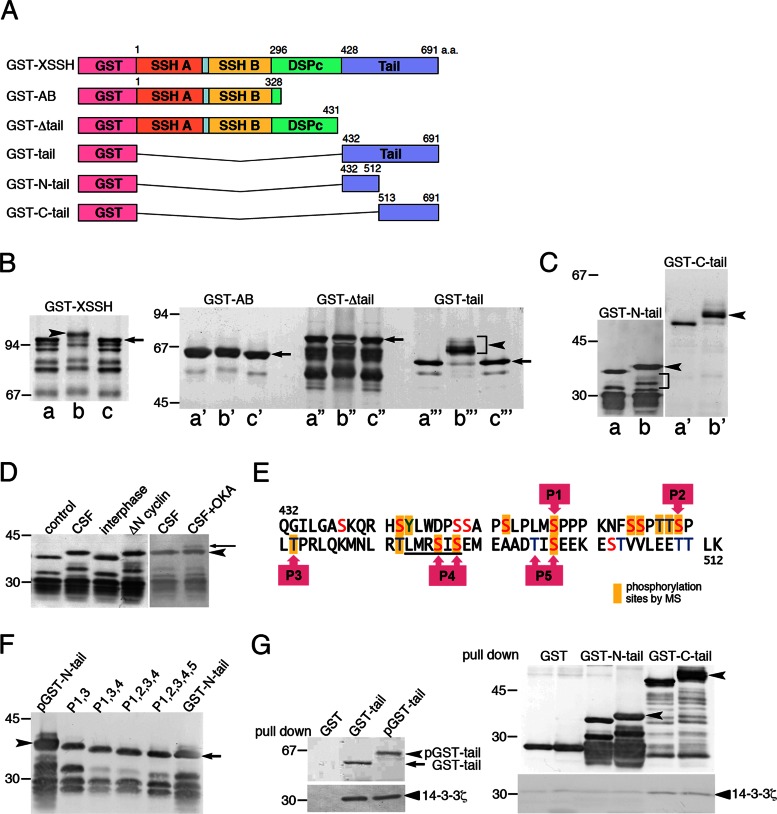
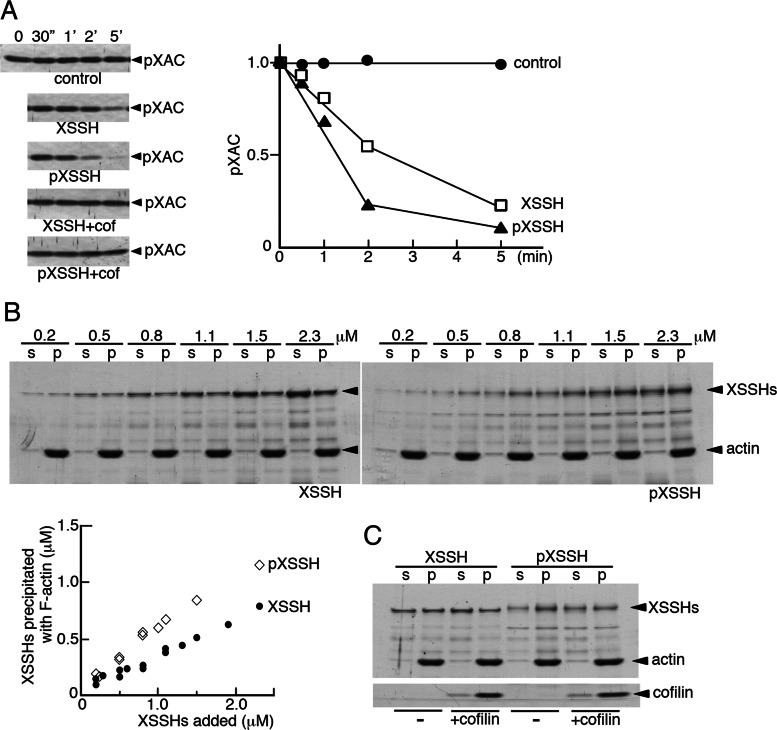
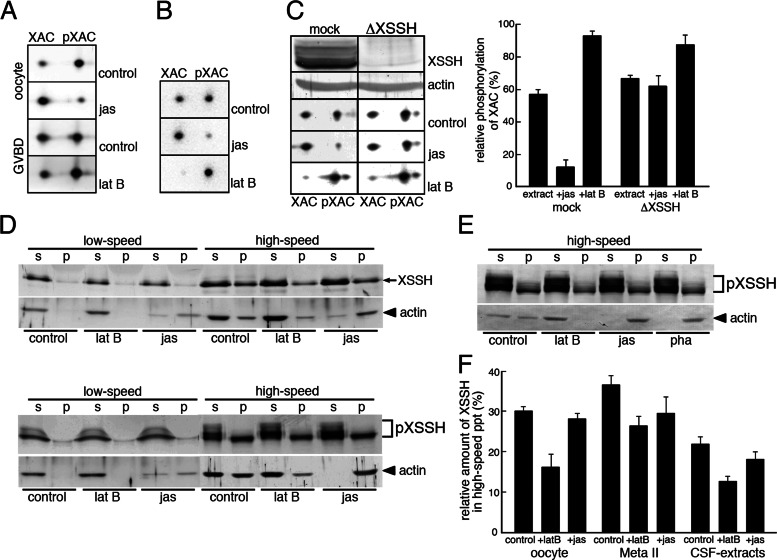
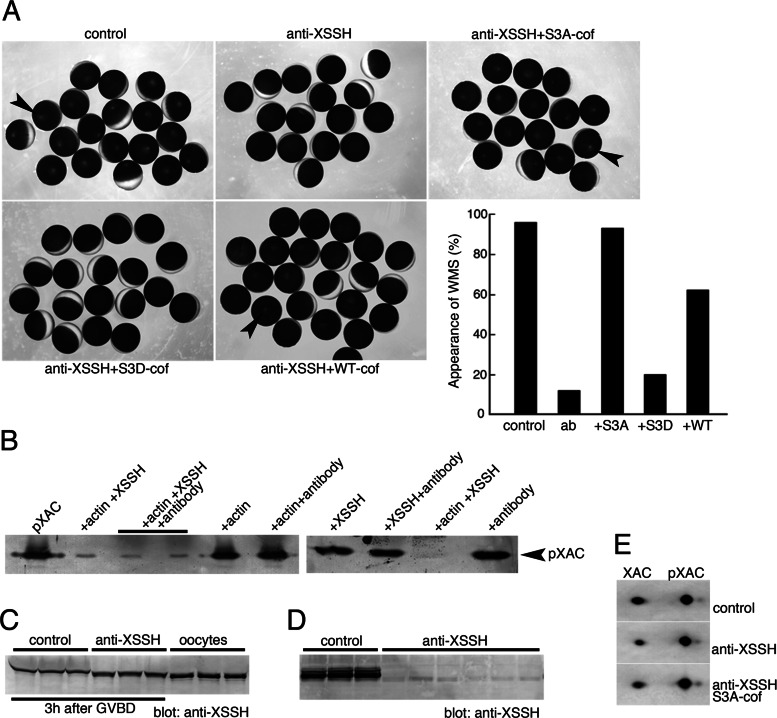
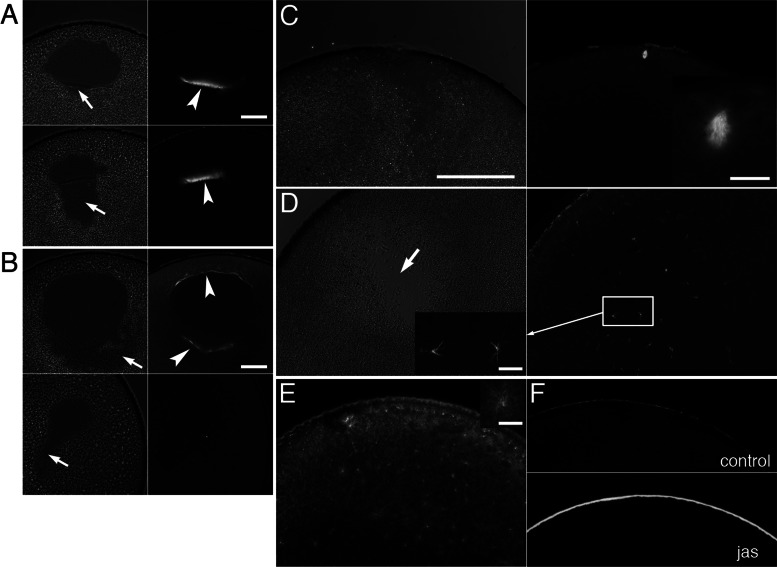
We identify Xenopus ADF/cofilin (XAC) and its activator, Slingshot phosphatase (XSSH), as key regulators of actin dynamics essential for spindle microtubule assembly during Xenopus oocyte maturation. Phosphorylation of XSSH at multiple sites within the tail domain occurs just after germinal vesicle breakdown (GVBD) and is accompanied by dephosphorylation of XAC, which was mostly phosphorylated in immature oocytes. This XAC dephosphorylation after GVBD is completely suppressed by latrunculin B, an actin monomer-sequestering drug. On the other hand, jasplakinolide, an F-actin-stabilizing drug, induces dephosphorylation of XAC. Effects of latrunculin B and jasplakinolide are reconstituted in cytostatic factor-arrested extracts (CSF extracts), and XAC dephosphorylation is abolished by depletion of XSSH from CSF extracts, suggesting that XSSH functions as an actin filament sensor to facilitate actin filament dynamics via XAC activation. Injection of anti-XSSH antibody, which blocks full phosphorylation of XSSH after GVBD, inhibits both meiotic spindle formation and XAC dephosphorylation. Coinjection of constitutively active XAC with the antibody suppresses this phenotype. Treatment of oocytes with jasplakinolide also impairs spindle formation. These results strongly suggest that elevation of actin dynamics by XAC activation through XSSH phosphorylation is required for meiotic spindle assembly in Xenopus laevis.
Figures
Similar articles
-
Reorganization of actin filaments by ADF/cofilin is involved in formation of microtubule structures during Xenopus oocyte maturation.Mol Biol Cell. 2015 Dec 1;26(24):4387-400. doi: 10.1091/mbc.E15-01-0035. Epub 2015 Sep 30. Mol Biol Cell. 2015. PMID: 26424802 Free PMC article.
-
Involvement of slingshot in the Rho-mediated dephosphorylation of ADF/cofilin during Xenopus cleavage.Zoolog Sci. 2005 Sep;22(9):971-84. doi: 10.2108/zsj.22.971. Zoolog Sci. 2005. PMID: 16219977
-
Functional involvement of Xenopus homologue of ADF/cofilin phosphatase, slingshot (XSSH), in the gastrulation movement.Zoolog Sci. 2005 Sep;22(9):955-69. doi: 10.2108/zsj.22.955. Zoolog Sci. 2005. PMID: 16219976
-
Signaling mechanisms and functional roles of cofilin phosphorylation and dephosphorylation.Cell Signal. 2013 Feb;25(2):457-69. doi: 10.1016/j.cellsig.2012.11.001. Epub 2012 Nov 12. Cell Signal. 2013. PMID: 23153585 Review.
-
Cofilin phosphatases and regulation of actin dynamics.Curr Opin Cell Biol. 2006 Feb;18(1):26-31. doi: 10.1016/j.ceb.2005.11.005. Epub 2005 Dec 7. Curr Opin Cell Biol. 2006. PMID: 16337782 Review.
Cited by
-
Reorganization of actin filaments by ADF/cofilin is involved in formation of microtubule structures during Xenopus oocyte maturation.Mol Biol Cell. 2015 Dec 1;26(24):4387-400. doi: 10.1091/mbc.E15-01-0035. Epub 2015 Sep 30. Mol Biol Cell. 2015. PMID: 26424802 Free PMC article.
-
Roles of actin binding proteins in mammalian oocyte maturation and beyond.Cell Cycle. 2016 Jul 17;15(14):1830-43. doi: 10.1080/15384101.2016.1181239. Epub 2016 May 6. Cell Cycle. 2016. PMID: 27152960 Free PMC article. Review.
-
The subcortical maternal complex controls symmetric division of mouse zygotes by regulating F-actin dynamics.Nat Commun. 2014 Sep 11;5:4887. doi: 10.1038/ncomms5887. Nat Commun. 2014. PMID: 25208553 Free PMC article.
References
-
- Arber S, Barbayannis FA, Hanser H, Schneider C, Stanyon CA, Bernard O, Caroni P. Regulation of actin dynamics through phosphorylation of cofilin by LIM-kinase. Nature. 1998;393:805–809. - PubMed
-
- Azoury J, Lee KW, Georget V, Hikal P, Verlhac MH. Symmetry breaking in mouse oocytes requires transient F-actin meshwork destabilization. Development. 2011;138:2903–2908. - PubMed
-
- Azoury J, Lee KW, Georget V, Rassinier P, Leader B, Verlhac MH. Spindle positioning in mouse oocytes relies on a dynamic meshwork of actin filaments. Curr Biol. 2008;18:1514–1519. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
