

Short-term depression of external globus pallidus-subthalamic nucleus synaptic transmission and implications for patterning subthalamic activity
- PMID: 23616523
- PMCID: PMC3678728
- DOI: 10.1523/JNEUROSCI.3576-12.2013
Short-term depression of external globus pallidus-subthalamic nucleus synaptic transmission and implications for patterning subthalamic activity
Abstract
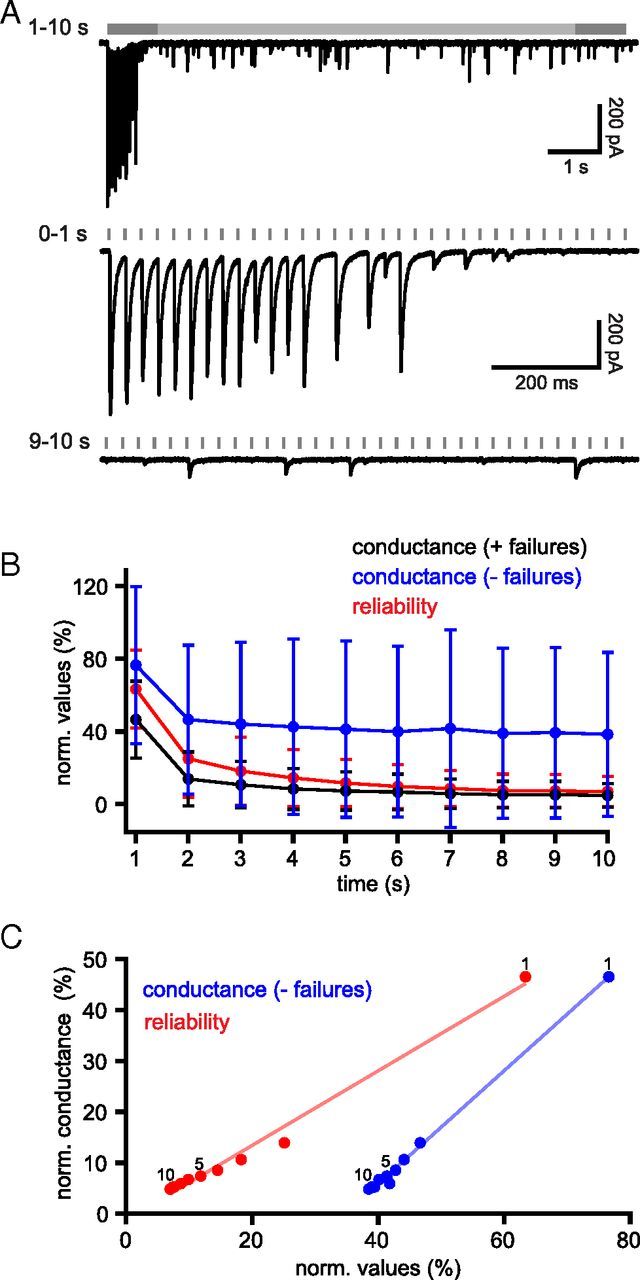
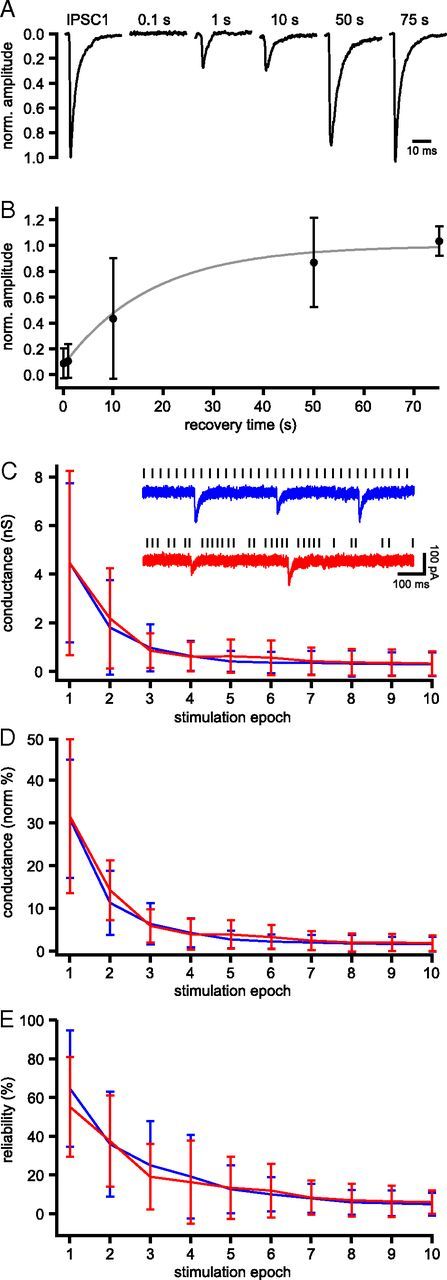
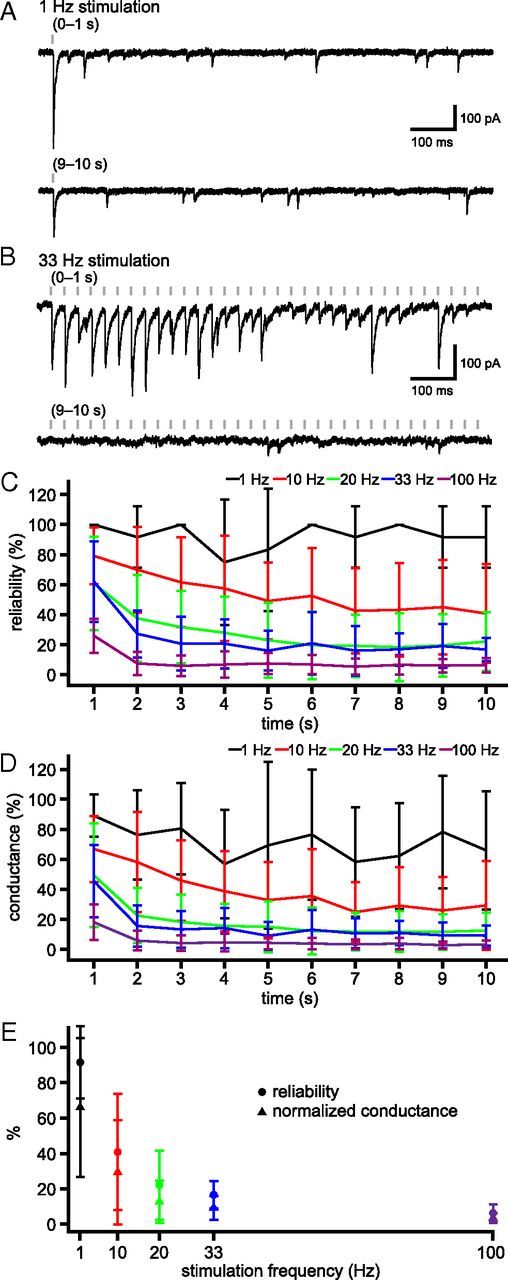
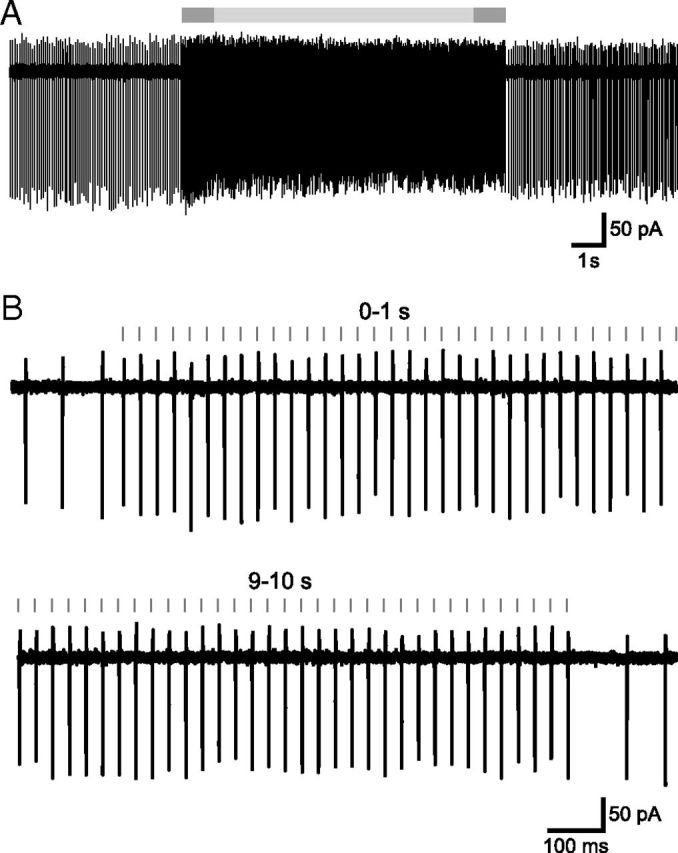
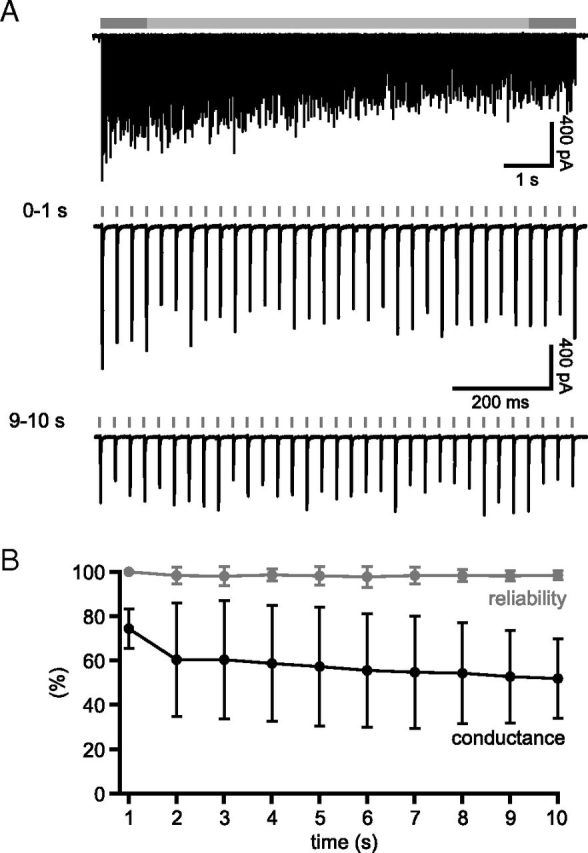
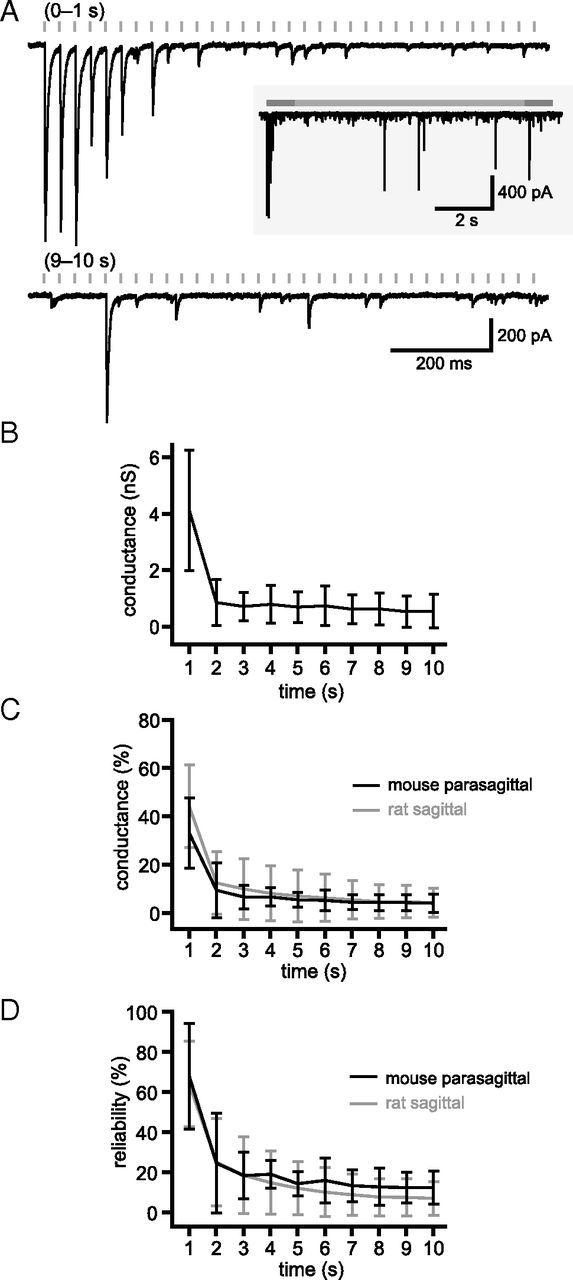
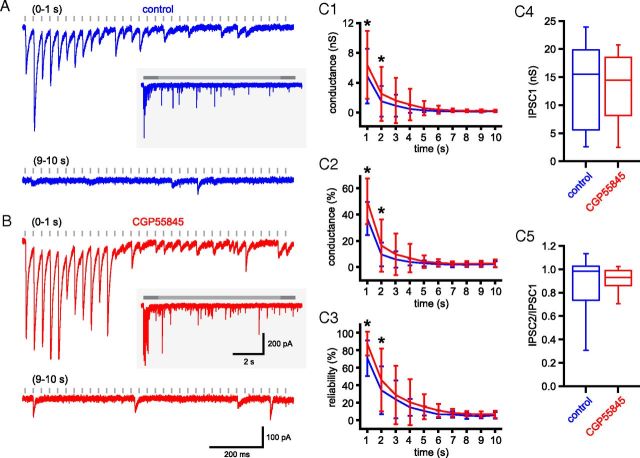
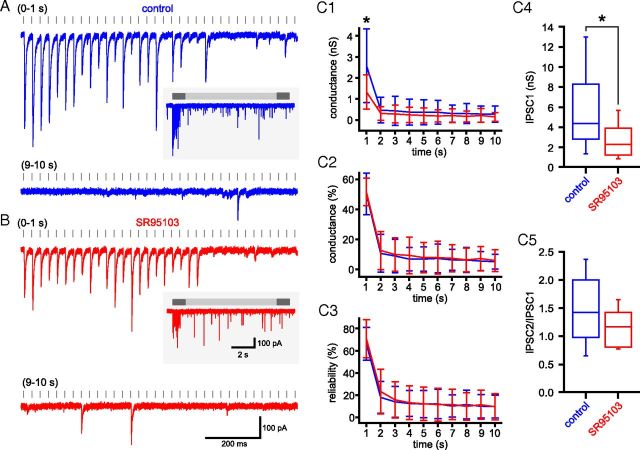
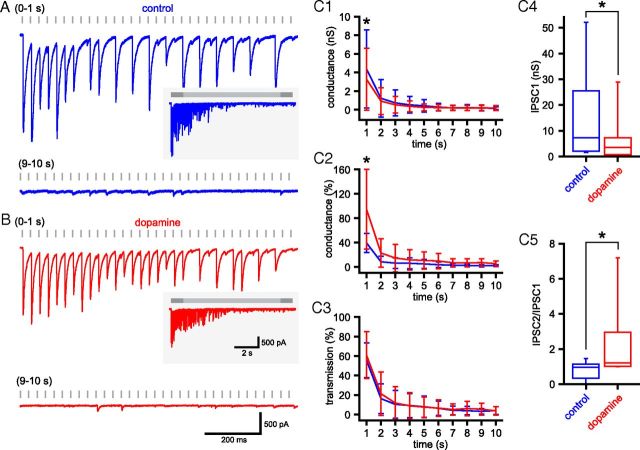
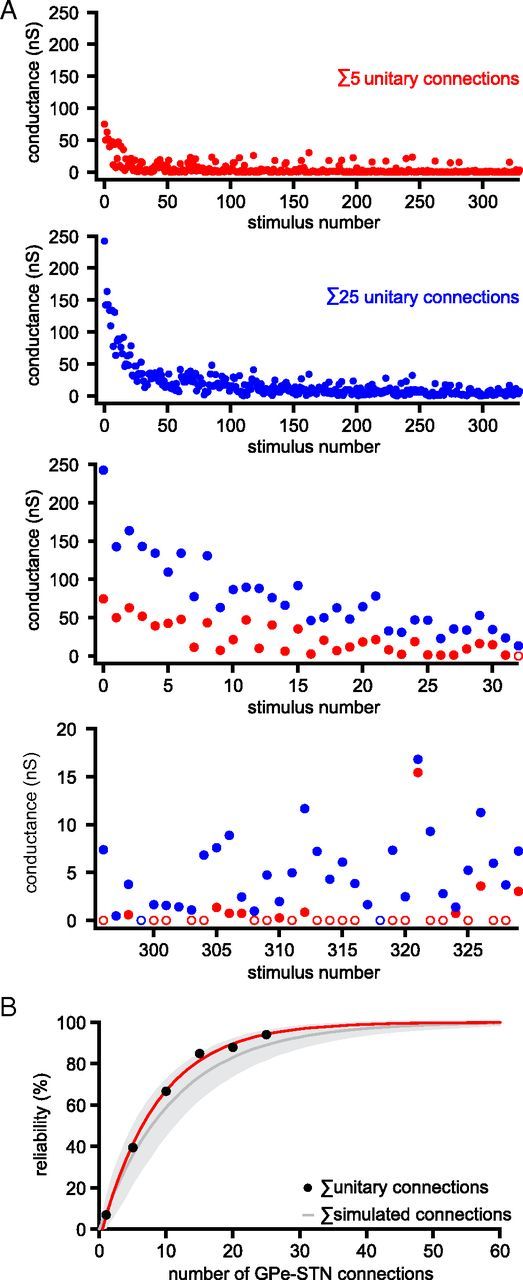
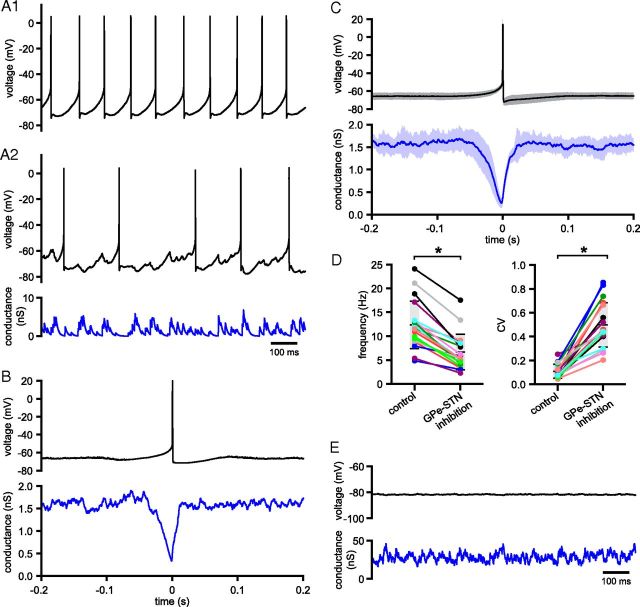
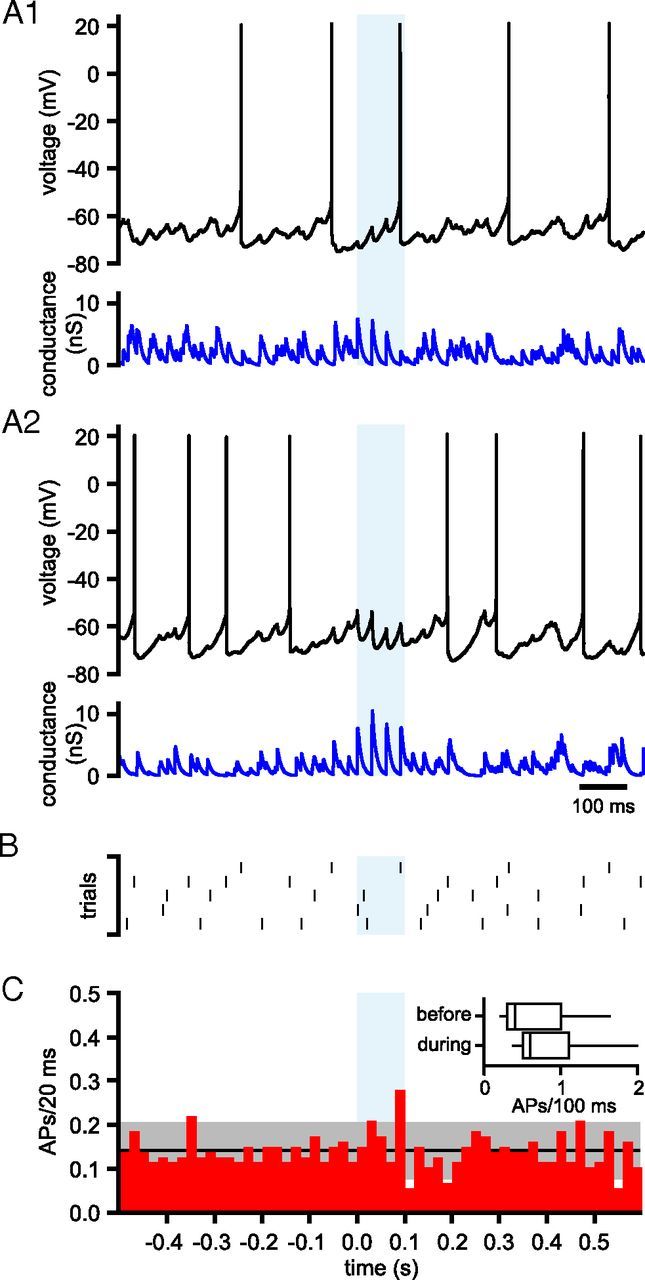
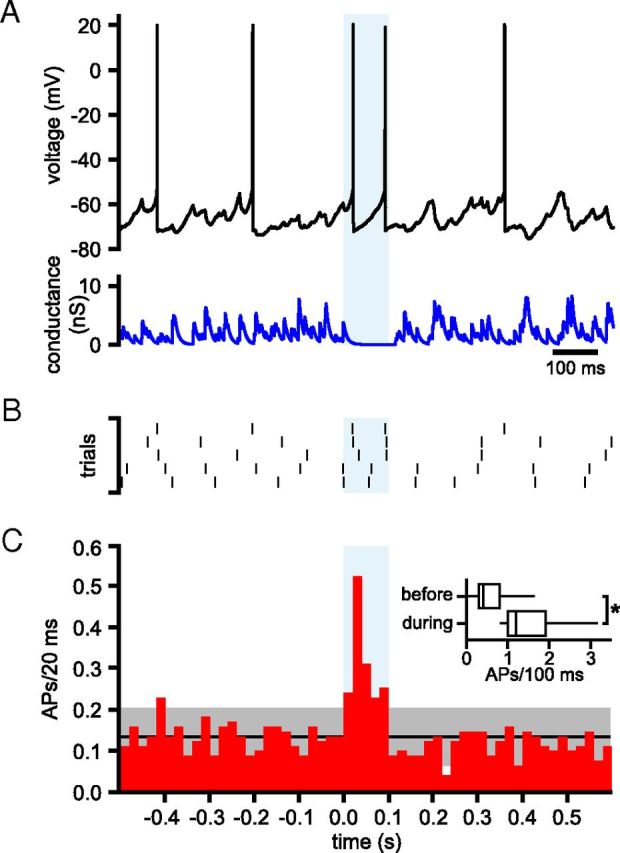
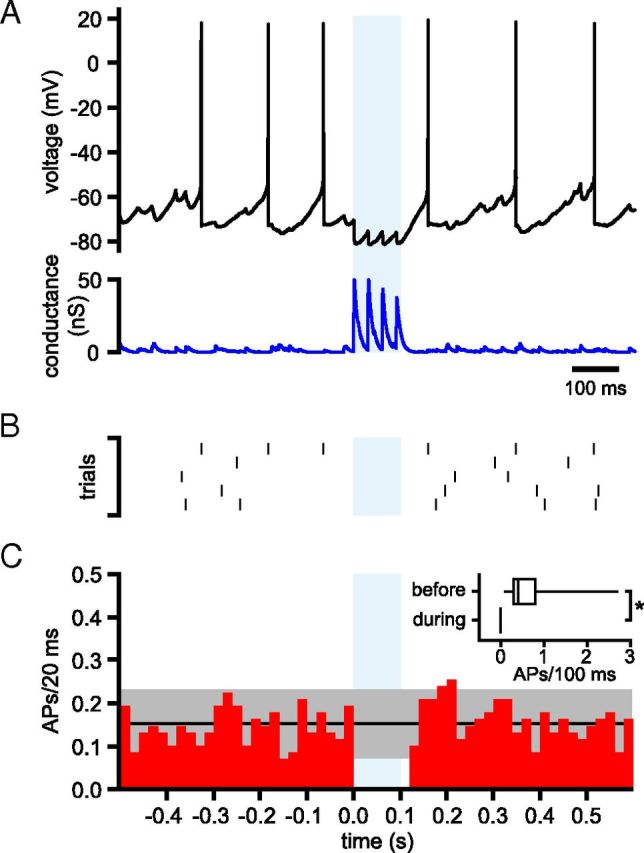
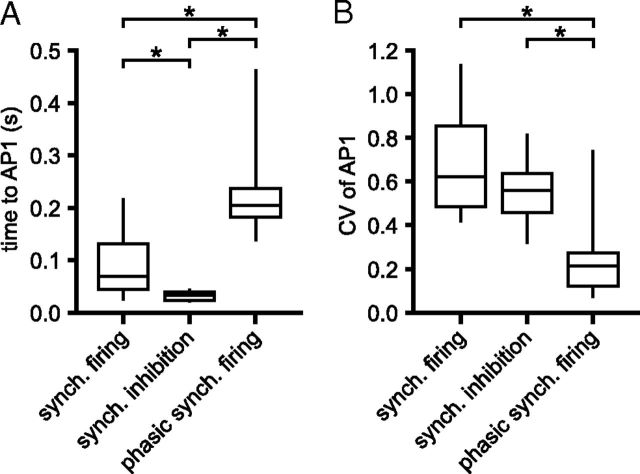
The frequency and pattern of activity in the reciprocally connected GABAergic external globus pallidus (GPe) and glutamatergic subthalamic nucleus (STN) are closely related to motor function. Although phasic, unitary GPe-STN inputs powerfully pattern STN activity ex vivo, correlated GPe-STN activity is not normally observed in vivo. To test the hypothesis that the GPe's influence is constrained by short-term synaptic depression, unitary GPe-STN inputs were stimulated in rat and mouse brain slices at rates and in patterns that mimicked GPe activity in vivo. Together with connectivity estimates these data were then used to simulate GPe-STN transmission. Unitary GPe-STN synaptic connections initially generated large conductances and transmitted reliably. However, the amplitude and reliability of transmission declined rapidly (τ = 0.6 ± 0.5 s) to <10% of their initial values when connections were stimulated at the mean rate of GPe activity in vivo (33 Hz). Recovery from depression (τ = 17.3 ± 18.9 s) was also longer than pauses in tonic GPe activity in vivo. Depression was the result of the limited supply of release-ready vesicles and was in sharp contrast to Calyx of Held transmission, which exhibited 100% reliability. Injection of simulated GPe-STN conductances revealed that synaptic depression caused tonic, nonsynchronized GPe-STN activity to disrupt rather than abolish autonomous STN activity. Furthermore, synchronous inhibition of tonically active GPe-STN neurons or phasic activity of GPe-STN neurons reliably patterned STN activity through disinhibition and inhibition, respectively. Together, these data argue that the frequency and pattern of GPe activity profoundly influence its transmission to the STN.
Figures
References
-
- Atherton JF, Kitano K, Baufreton J, Fan K, Wokosin D, Tkatch T, Shigemoto R, Surmeier DJ, Bevan MD. Selective participation of somatodendritic HCN channels in inhibitory but not excitatory synaptic integration in neurons of the subthalamic nucleus. J Neurosci. 2010;30:16025–16040. doi: 10.1523/JNEUROSCI.3898-10.2010. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources