

EBE, an AP2/ERF transcription factor highly expressed in proliferating cells, affects shoot architecture in Arabidopsis
- PMID: 23616605
- PMCID: PMC3668074
- DOI: 10.1104/pp.113.214049
EBE, an AP2/ERF transcription factor highly expressed in proliferating cells, affects shoot architecture in Arabidopsis
Abstract
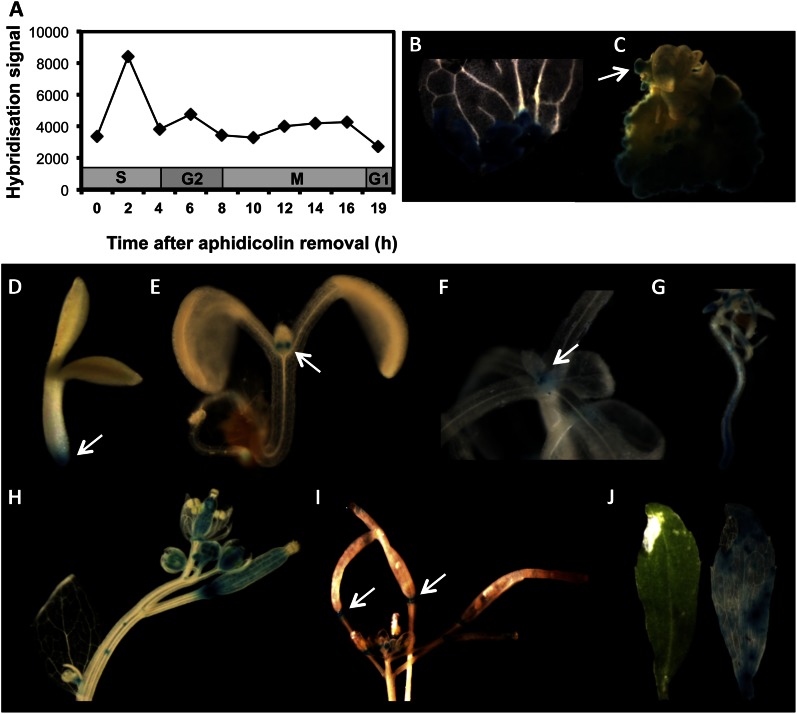
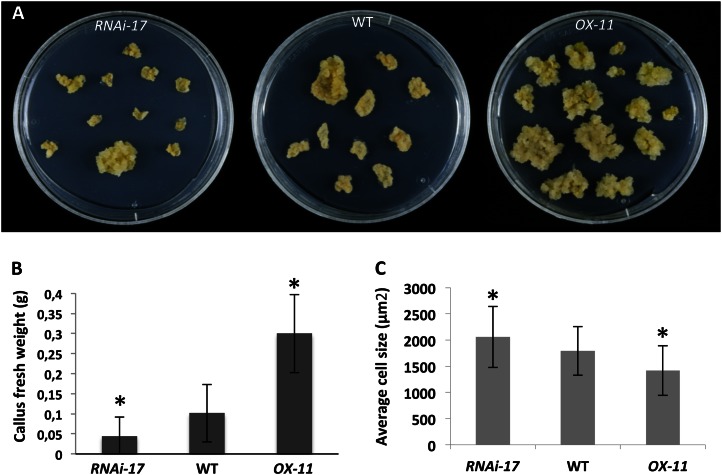
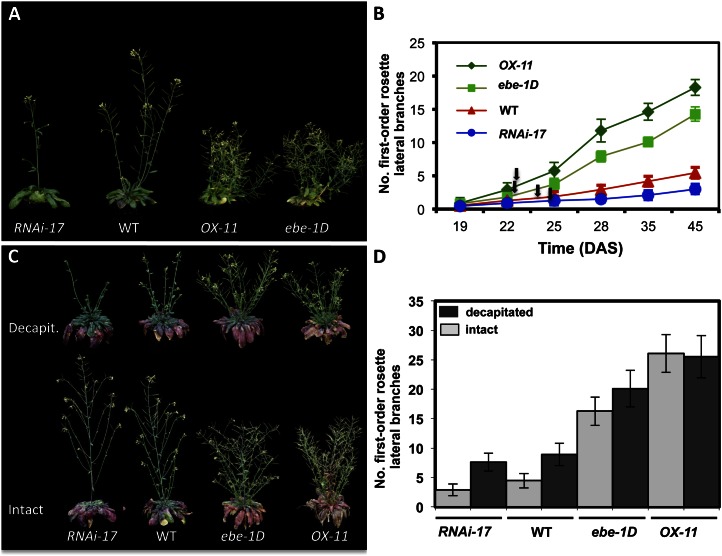
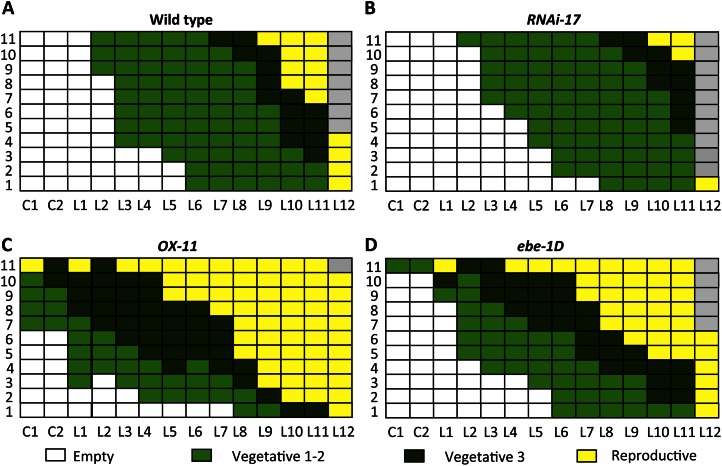
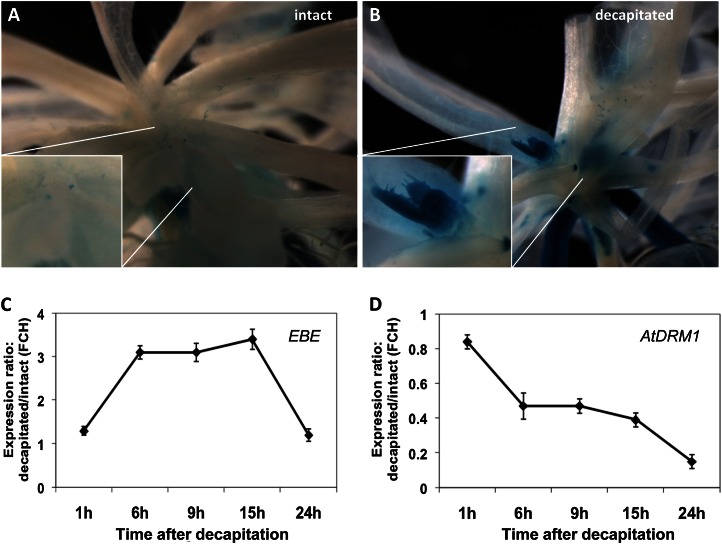
We report about ERF BUD ENHANCER (EBE; At5g61890), a transcription factor that affects cell proliferation as well as axillary bud outgrowth and shoot branching in Arabidopsis (Arabidopsis thaliana). EBE encodes a member of the APETALA2/ETHYLENE RESPONSE FACTOR (AP2/ERF) transcription factor superfamily; the gene is strongly expressed in proliferating cells and is rapidly and transiently up-regulated in axillary meristems upon main stem decapitation. Overexpression of EBE promotes cell proliferation in growing calli, while the opposite is observed in EBE-RNAi lines. EBE overexpression also stimulates axillary bud formation and outgrowth, while repressing it results in inhibition of bud growth. Global transcriptome analysis of estradiol-inducible EBE overexpression lines revealed 48 EBE early-responsive genes, of which 14 were up-regulated and 34 were down-regulated. EBE activates several genes involved in cell cycle regulation and dormancy breaking, including D-type cyclin CYCD3;3, transcription regulator DPa, and BRCA1-ASSOCIATED RING DOMAIN1. Among the down-regulated genes were DORMANCY-ASSOCIATED PROTEIN1 (AtDRM1), AtDRM1 homolog, MEDIATOR OF ABA-REGULATED DORMANCY1, and ZINC FINGER HOMEODOMAIN5. Our data indicate that the effect of EBE on shoot branching likely results from an activation of genes involved in cell cycle regulation and dormancy breaking.
Figures

References
-
- Aggarwal P, Padmanabhan B, Bhat A, Sarvepalli K, Sadhale PP, Nath U. (2011) The TCP4 transcription factor of Arabidopsis blocks cell division in yeast at G1→S transition. Biochem Biophys Res Commun 410: 276–281 - PubMed
-
- Bakan B, Hamberg M, Perrocheau L, Maume D, Rogniaux H, Tranquet O, Rondeau C, Blein J-P, Ponchet M, Marion D. (2006) Specific adduction of plant lipid transfer protein by an allene oxide generated by 9-lipoxygenase and allene oxide synthase. J Biol Chem 281: 38981–38988 - PubMed
-
- Balazadeh S, Riaño-Pachón DM, Mueller-Roeber B. (2008) Transcription factors regulating leaf senescence in Arabidopsis thaliana. Plant Biol (Stuttg) (Suppl 1) 10: 63–75 - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
