

TERRA, hnRNP A1, and DNA-PKcs Interactions at Human Telomeres
- PMID: 23616949
- PMCID: PMC3628365
- DOI: 10.3389/fonc.2013.00091
TERRA, hnRNP A1, and DNA-PKcs Interactions at Human Telomeres
Abstract
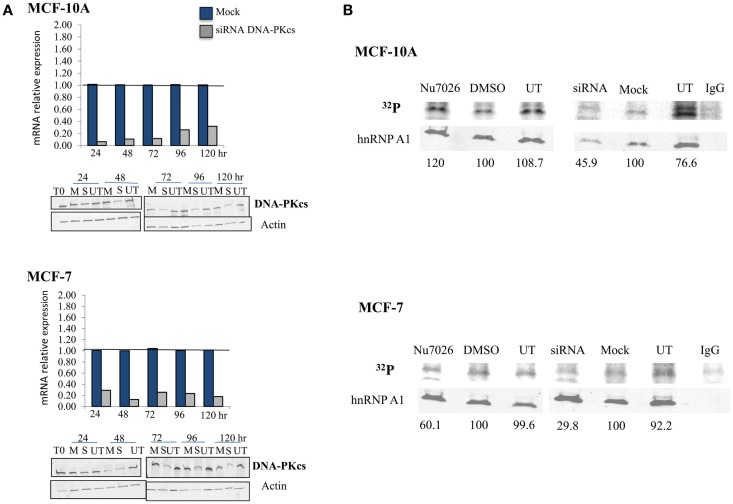
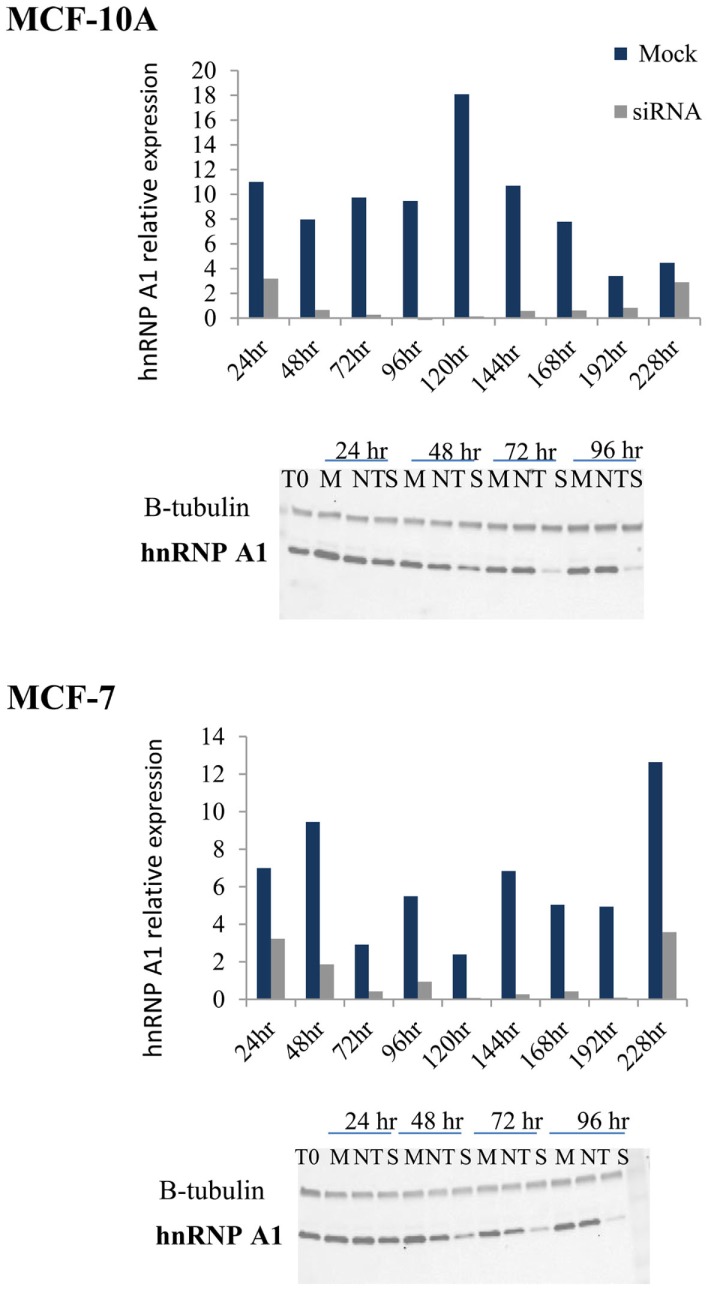
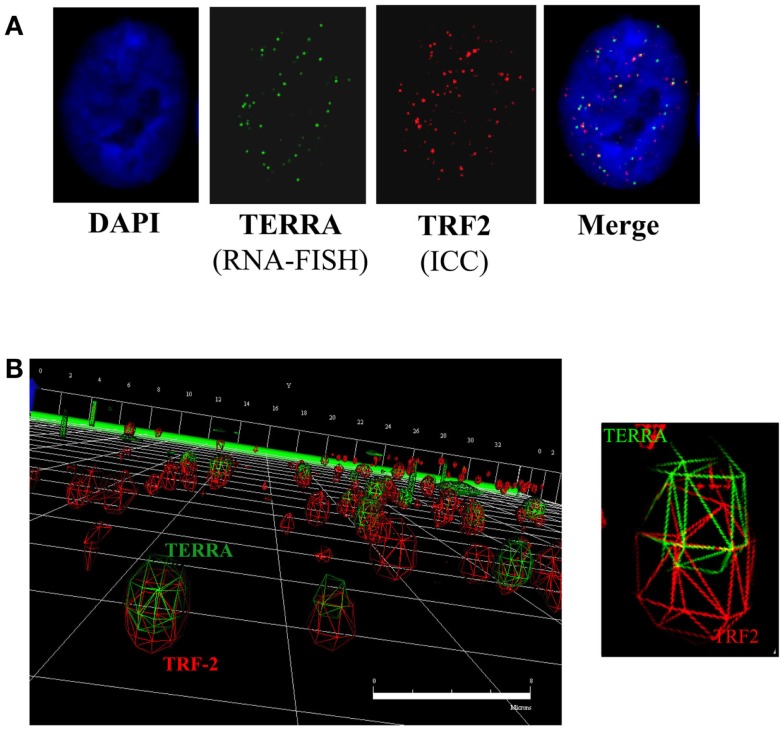
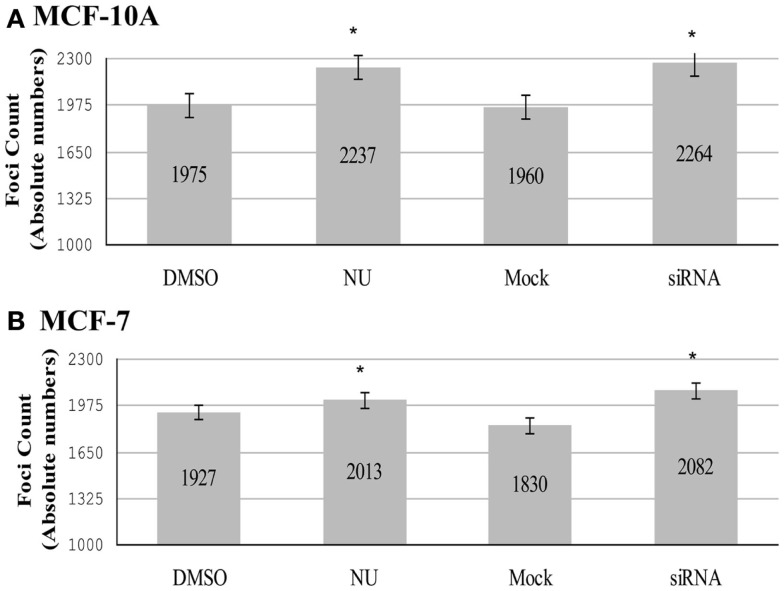
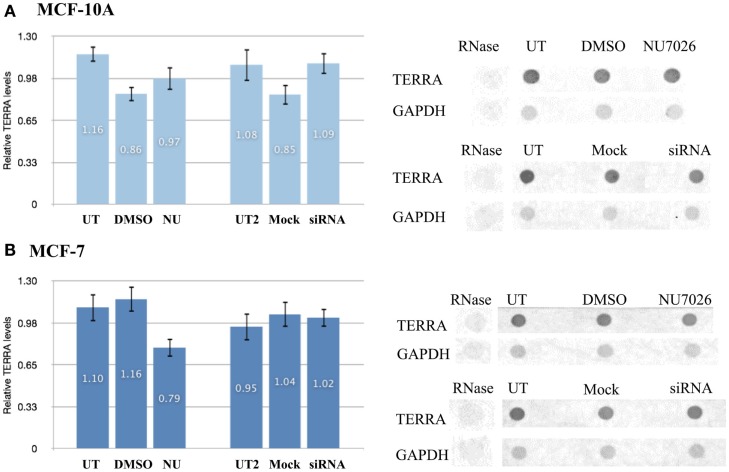
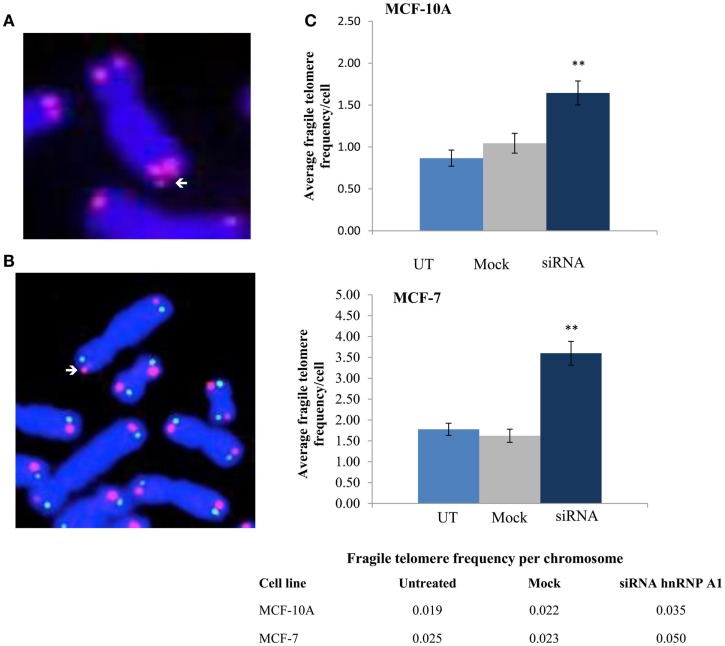
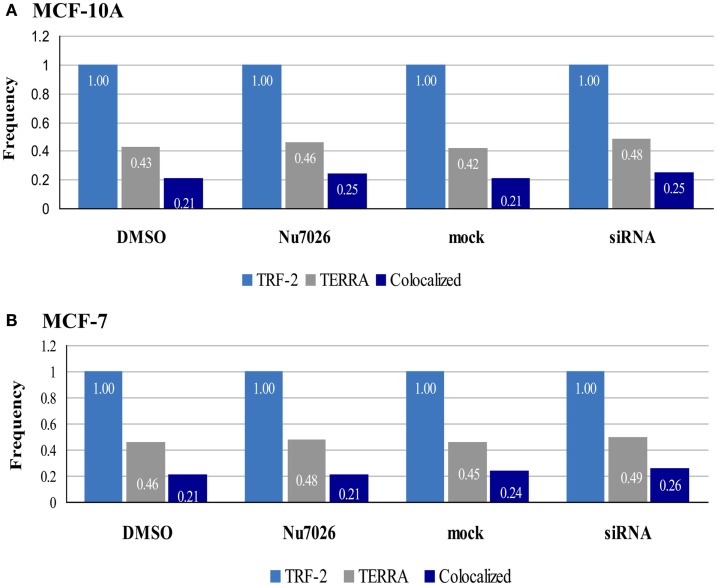
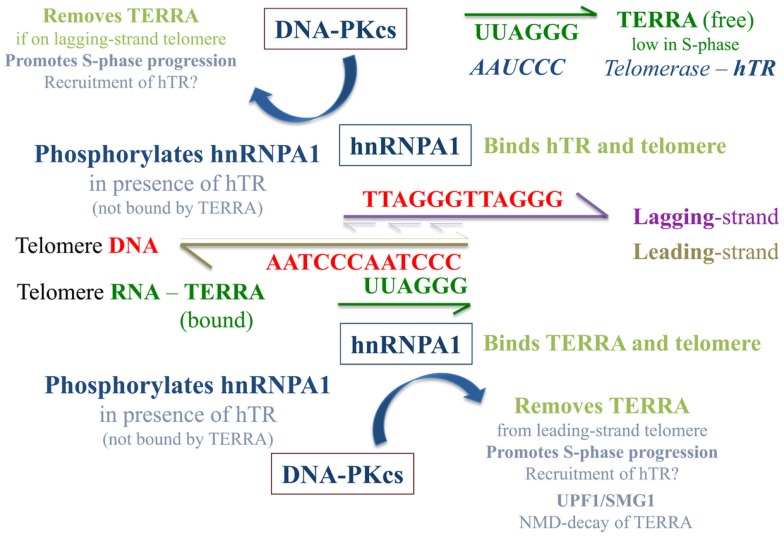
Maintenance of telomeres, repetitive elements at eukaryotic chromosomal termini, and the end-capping structure and function they provide, are imperative for preserving genome integrity and stability. The discovery that telomeres are transcribed into telomere repeat containing RNA (TERRA) has revolutionized our view of this repetitive, rather unappreciated region of the genome. We have previously shown that the non-homologous end-joining, shelterin associated DNA dependent protein kinase catalytic subunit (DNA-PKcs) participates in mammalian telomeric end-capping, exclusively at telomeres created by leading-strand synthesis. Here, we explore potential roles of DNA-PKcs and its phosphorylation target heterogeneous nuclear ribonucleoprotein A1 (hnRNP A1) in the localization of TERRA at human telomeres. Evaluation of co-localized foci utilizing RNA-FISH and three-dimensional (3D) reconstruction strategies provided evidence that both inhibition of DNA-PKcs kinase activity and siRNA depletion of hnRNP A1 result in accumulation of TERRA at individual telomeres; depletion of hnRNP A1 also resulted in increased frequencies of fragile telomeres. These observations are consistent with previous demonstrations that decreased levels of the nonsense RNA-mediated decay factors SMG1 and UPF1 increase TERRA at telomeres and interfere with replication of leading-strand telomeres. We propose that hTR mediated stimulation of DNA-PKcs and subsequent phosphorylation of hnRNP A1 influences the cell cycle dependent distribution of TERRA at telomeres by contributing to the removal of TERRA from telomeres, an action important for progression of S-phase, and thereby facilitating efficient telomere replication and end-capping.
Keywords: DNA-PKcs; TERRA; hTR; hnRNP A1; strand-specificity; telomeres.
Figures

Similar articles
-
Phase separation of hnRNPA1 and TERRA regulates telomeric stability.J Mol Cell Biol. 2025 Mar 21;16(9):mjae037. doi: 10.1093/jmcb/mjae037. J Mol Cell Biol. 2025. PMID: 39313323 Free PMC article.
-
DNA-PKcs phosphorylates hnRNP-A1 to facilitate the RPA-to-POT1 switch and telomere capping after replication.Nucleic Acids Res. 2015 Jul 13;43(12):5971-83. doi: 10.1093/nar/gkv539. Epub 2015 May 20. Nucleic Acids Res. 2015. PMID: 25999341 Free PMC article.
-
Protein Phosphatase 2A-Dependent Mitotic hnRNPA1 Dephosphorylation and TERRA Formation Facilitate Telomere Capping.Mol Cancer Res. 2022 Apr 1;20(4):583-595. doi: 10.1158/1541-7786.MCR-21-0581. Mol Cancer Res. 2022. PMID: 34933911
-
DNA-dependent protein kinase in telomere maintenance and protection.Cell Mol Biol Lett. 2020 Jan 17;25:2. doi: 10.1186/s11658-020-0199-0. eCollection 2020. Cell Mol Biol Lett. 2020. PMID: 31988640 Free PMC article. Review.
-
Telomeric repeat-containing RNA TERRA: a noncoding RNA connecting telomere biology to genome integrity.Front Genet. 2015 Apr 14;6:143. doi: 10.3389/fgene.2015.00143. eCollection 2015. Front Genet. 2015. PMID: 25926849 Free PMC article. Review.
Cited by
-
Telomere Fragility and MiDAS: Managing the Gaps at the End of the Road.Genes (Basel). 2023 Jan 29;14(2):348. doi: 10.3390/genes14020348. Genes (Basel). 2023. PMID: 36833275 Free PMC article. Review.
-
Heterogeneous Nuclear Ribonucleoproteins Involved in the Functioning of Telomeres in Malignant Cells.Int J Mol Sci. 2019 Feb 10;20(3):745. doi: 10.3390/ijms20030745. Int J Mol Sci. 2019. PMID: 30744200 Free PMC article. Review.
-
NHP2 downregulation counteracts hTR-mediated activation of the DNA damage response at ALT telomeres.EMBO J. 2021 Mar 15;40(6):e106336. doi: 10.15252/embj.2020106336. Epub 2021 Feb 17. EMBO J. 2021. PMID: 33595114 Free PMC article.
-
Phase separation of hnRNPA1 and TERRA regulates telomeric stability.J Mol Cell Biol. 2025 Mar 21;16(9):mjae037. doi: 10.1093/jmcb/mjae037. J Mol Cell Biol. 2025. PMID: 39313323 Free PMC article.
-
IN-DEPTH MAPPING OF DNA-PKcs SIGNALING UNCOVERS CONSERVED FEATURES OF ITS KINASE SPECIFICITY.bioRxiv [Preprint]. 2024 Jan 19:2024.01.17.576037. doi: 10.1101/2024.01.17.576037. bioRxiv. 2024. Update in: J Biol Chem. 2024 Aug;300(8):107513. doi: 10.1016/j.jbc.2024.107513. PMID: 38293078 Free PMC article. Updated. Preprint.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources