

Histidine biosynthesis, its regulation and biotechnological application in Corynebacterium glutamicum
- PMID: 23617600
- PMCID: PMC3896937
- DOI: 10.1111/1751-7915.12055
Histidine biosynthesis, its regulation and biotechnological application in Corynebacterium glutamicum
Abstract
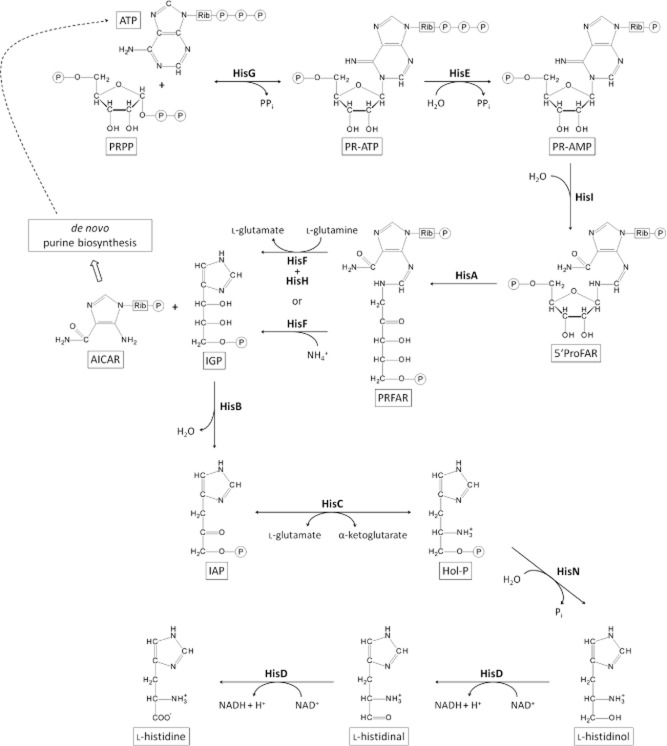
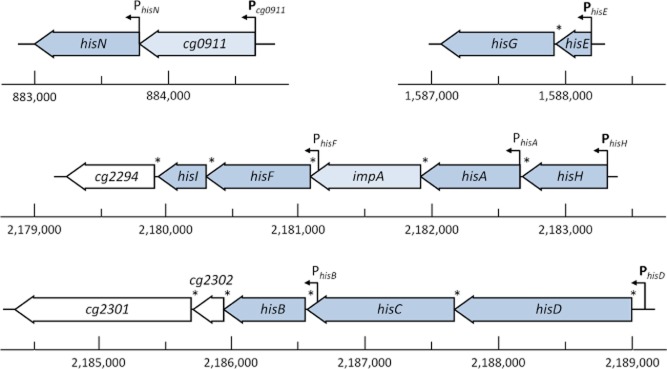
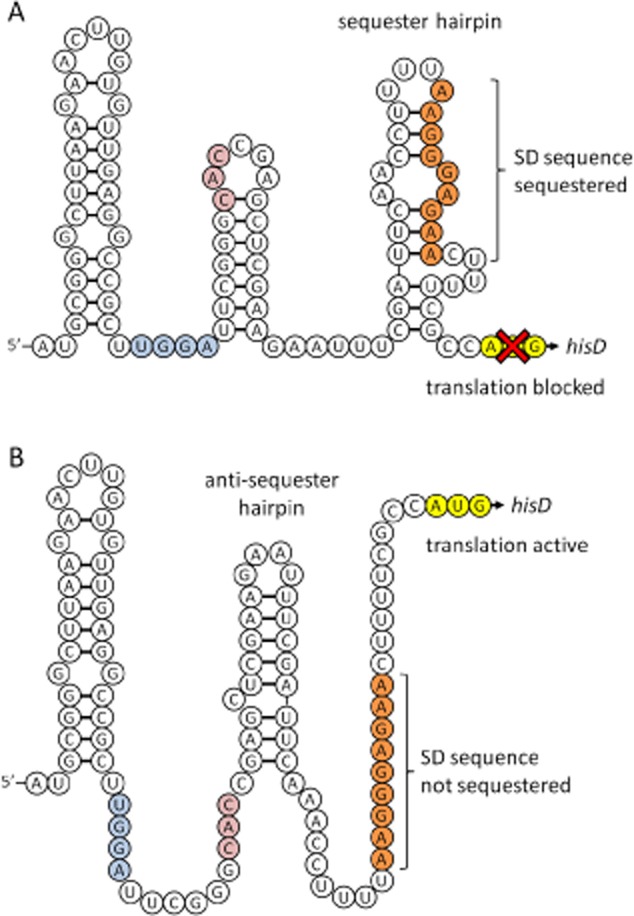
l-Histidine biosynthesis is an ancient metabolic pathway present in bacteria, archaea, lower eukaryotes, and plants. For decades l-histidine biosynthesis has been studied mainly in Escherichia coli and Salmonella typhimurium, revealing fundamental regulatory processes in bacteria. Furthermore, in the last 15 years this pathway has been also investigated intensively in the industrial amino acid-producing bacterium Corynebacterium glutamicum, revealing similarities to E. coli and S. typhimurium, as well as differences. This review summarizes the current knowledge of l-histidine biosynthesis in C. glutamicum. The genes involved and corresponding enzymes are described, in particular focusing on the imidazoleglycerol-phosphate synthase (HisFH) and the histidinol-phosphate phosphatase (HisN). The transcriptional organization of his genes in C. glutamicum is also reported, including the four histidine operons and their promoters. Knowledge of transcriptional regulation during stringent response and by histidine itself is summarized and a translational regulation mechanism is discussed, as well as clues about a histidine transport system. Finally, we discuss the potential of using this knowledge to create or improve C. glutamicum strains for the industrial l-histidine production.
© 2013 The Authors. Microbial Biotechnology published by John Wiley & Sons Ltd and Society for Applied Microbiology.
Figures

References
-
- Adams E. The enzymatic synthesis of histidine from histidinol. J Biol Chem. 1954;209:829–846. - PubMed
-
- Alderwick LJ, Dover LG, Seidel M, Gande R, Sahm H, Eggeling L, Besra GS. Arabinan-deficient mutants of Corynebacterium glutamicum and the consequent flux in decaprenylmonophosphoryl-d-arabinose metabolism. Glycobiology. 2006;16:1073–1081. - PubMed
-
- Alifano P, Piscitelli C, Blasi V, Rivellini F, Nappo AG, Bruni CB, Carlomagno MS. Processing of a polycistronic mRNA requires a 5′ cis element and active translation. Mol Microbiol. 1992;6:787–798. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
