

Stabilization of an unusual salt bridge in ubiquitin by the extra C-terminal domain of the proteasome-associated deubiquitinase UCH37 as a mechanism of its exo specificity
- PMID: 23617878
- PMCID: PMC3898853
- DOI: 10.1021/bi4003106
Stabilization of an unusual salt bridge in ubiquitin by the extra C-terminal domain of the proteasome-associated deubiquitinase UCH37 as a mechanism of its exo specificity
Abstract
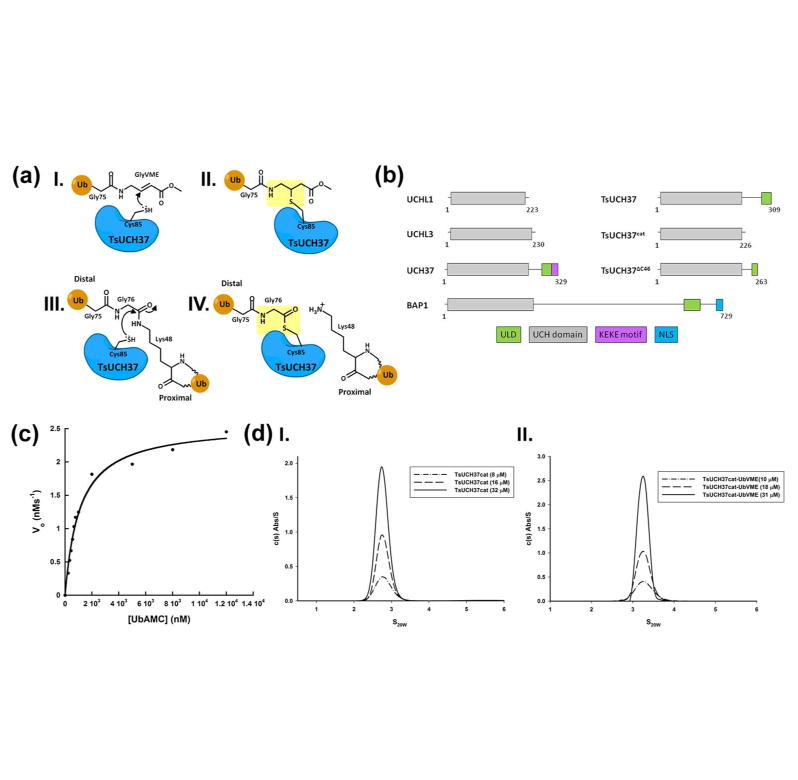
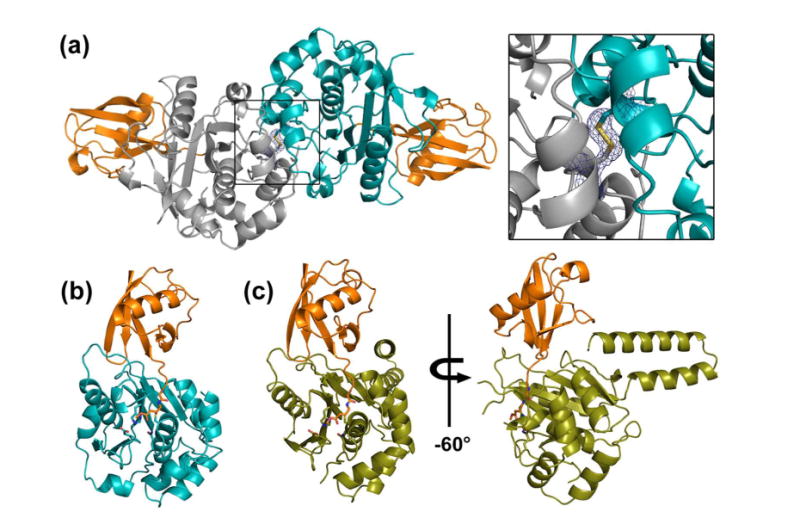
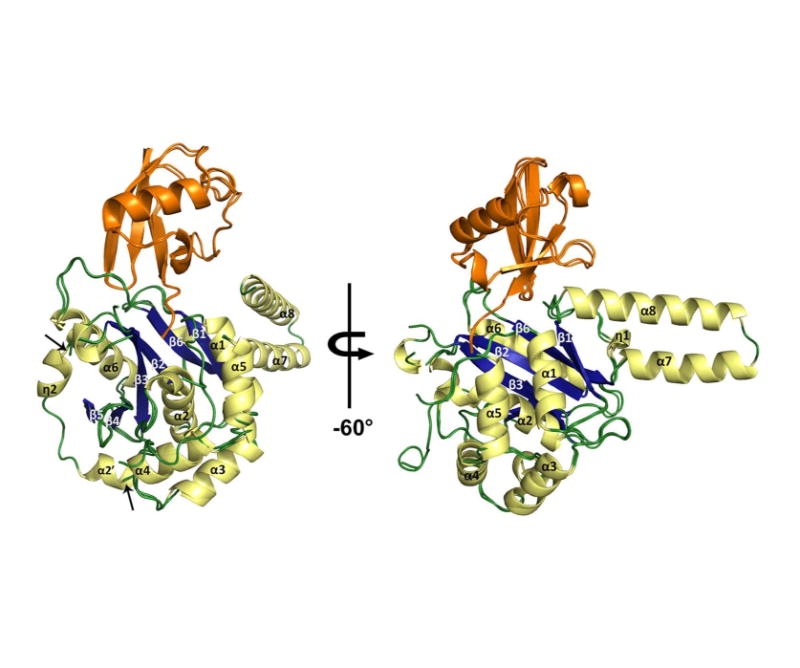
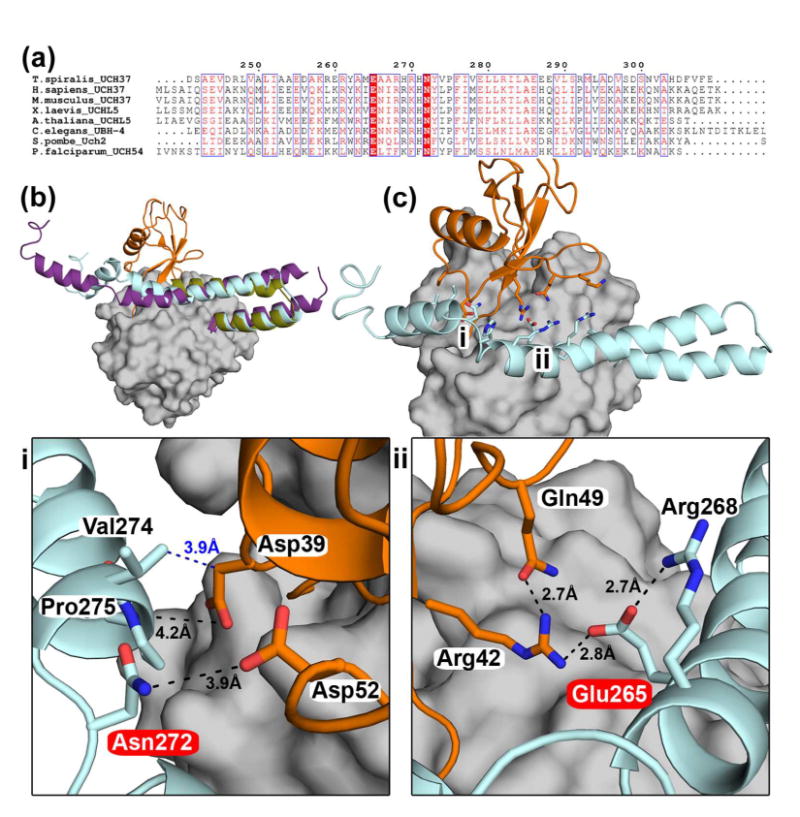
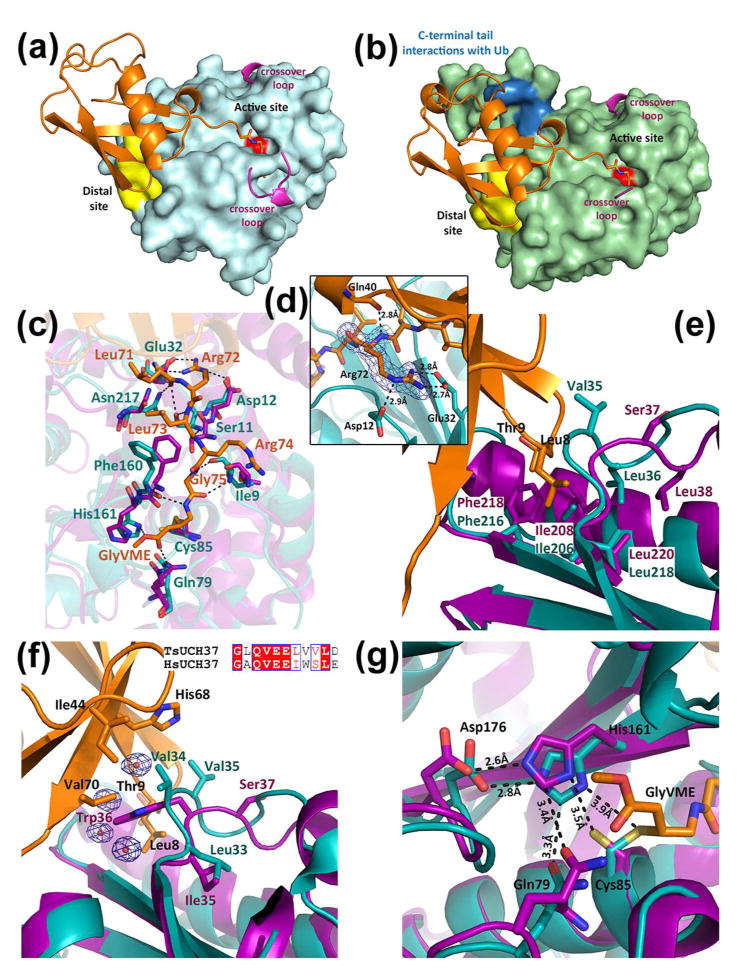
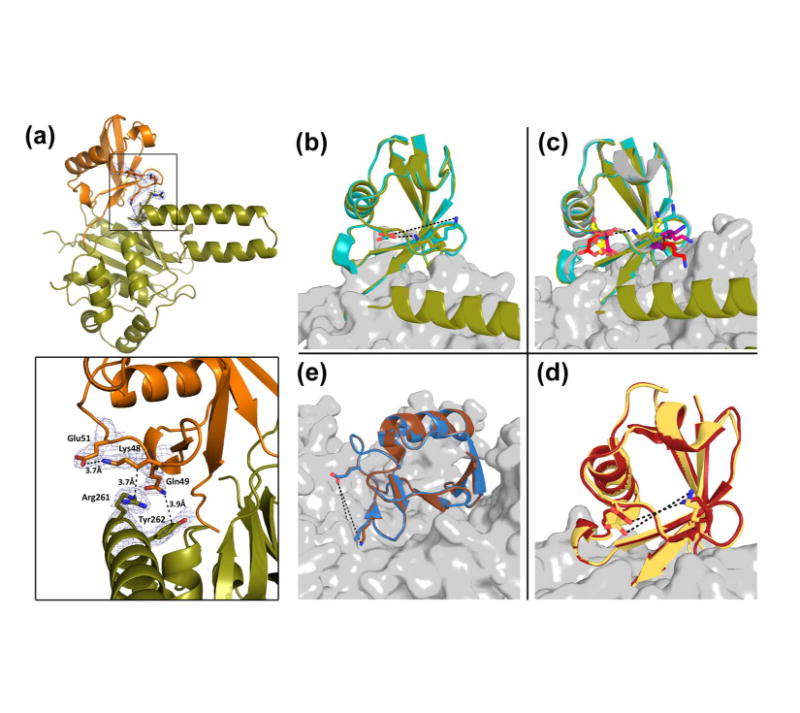
Ubiquitination is countered by a group of enzymes collectively called deubiquitinases (DUBs); ∼100 of them can be found in the human genome. One of the most interesting aspects of these enzymes is the ability of some members to selectively recognize specific linkage types between ubiquitin in polyubiquitin chains and their endo and exo specificity. The structural basis of exo-specific deubiquitination catalyzed by a DUB is poorly understood. UCH37, a cysteine DUB conserved from fungi to humans, is a proteasome-associated factor that regulates the proteasome by sequentially cleaving polyubiquitin chains from their distal ends, i.e., by exo-specific deubiquitination. In addition to the catalytic domain, the DUB features a functionally uncharacterized UCH37-like domain (ULD), presumed to keep the enzyme in an inhibited state in its proteasome-free form. Herein we report the crystal structure of two constructs of UCH37 from Trichinella spiralis in complex with a ubiquitin-based suicide inhibitor, ubiquitin vinyl methyl ester (UbVME). These structures show that the ULD makes direct contact with ubiquitin stabilizing a highly unusual intramolecular salt bridge between Lys48 and Glu51 of ubiquitin, an interaction that would be favored only with the distal ubiquitin but not with the internal ones in a Lys48-linked polyubiquitin chain. An inspection of 39 DUB-ubiquitin structures in the Protein Data Bank reveals the uniqueness of the salt bridge in ubiquitin bound to UCH37, an interaction that disappears when the ULD is deleted, as revealed in the structure of the catalytic domain alone bound to UbVME. The structural data are consistent with previously reported mutational data on the mammalian enzyme, which, together with the fact that the ULD residues that bind to ubiquitin are conserved, points to a similar mechanism behind the exo specificity of the human enzyme. To the best of our knowledge, these data provide the only structural example so far of how the exo specificity of a DUB can be determined by its noncatalytic domain. Thus, our data show that, contrary to its proposed inhibitory role, the ULD actually contributes to substrate recognition and could be a major determinant of the proteasome-associated function of UCH37. Moreover, our structures show that the unproductively oriented catalytic cysteine in the free enzyme is aligned correctly when ubiquitin binds, suggesting a mechanism for ubiquitin selectivity.
Figures

Similar articles
-
Branched ubiquitin chain binding and deubiquitination by UCH37 facilitate proteasome clearance of stress-induced inclusions.Elife. 2021 Nov 11;10:e72798. doi: 10.7554/eLife.72798. Elife. 2021. PMID: 34761751 Free PMC article.
-
Proteasome recruitment and activation of the Uch37 deubiquitinating enzyme by Adrm1.Nat Cell Biol. 2006 Sep;8(9):994-1002. doi: 10.1038/ncb1460. Epub 2006 Aug 13. Nat Cell Biol. 2006. PMID: 16906146
-
A ubiquitinome analysis to study the functional roles of the proteasome associated deubiquitinating enzymes USP14 and UCH37.J Proteomics. 2022 Jun 30;262:104592. doi: 10.1016/j.jprot.2022.104592. Epub 2022 Apr 27. J Proteomics. 2022. PMID: 35489684
-
The emerging role of Deubiquitinases (DUBs) in parasites: A foresight review.Front Cell Infect Microbiol. 2022 Sep 27;12:985178. doi: 10.3389/fcimb.2022.985178. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 36237424 Free PMC article. Review.
-
Reading the ubiquitin postal code.Curr Opin Struct Biol. 2011 Dec;21(6):792-801. doi: 10.1016/j.sbi.2011.09.009. Epub 2011 Oct 27. Curr Opin Struct Biol. 2011. PMID: 22036065 Review.
Cited by
-
Turnip yellow mosaic virus protease binds ubiquitin suboptimally to fine-tune its deubiquitinase activity.J Biol Chem. 2020 Oct 2;295(40):13769-13783. doi: 10.1074/jbc.RA120.014628. Epub 2020 Jul 30. J Biol Chem. 2020. PMID: 32732284 Free PMC article.
-
A deubiquitylase with an unusually high-affinity ubiquitin-binding domain from the scrub typhus pathogen Orientia tsutsugamushi.Nat Commun. 2020 May 11;11(1):2343. doi: 10.1038/s41467-020-15985-4. Nat Commun. 2020. PMID: 32393759 Free PMC article.
-
A Wolbachia deubiquitylating enzyme induces cytoplasmic incompatibility.Nat Microbiol. 2017 Mar 1;2:17007. doi: 10.1038/nmicrobiol.2017.7. Nat Microbiol. 2017. PMID: 28248294 Free PMC article.
-
The BAP1/ASXL2 Histone H2A Deubiquitinase Complex Regulates Cell Proliferation and Is Disrupted in Cancer.J Biol Chem. 2015 Nov 27;290(48):28643-63. doi: 10.1074/jbc.M115.661553. Epub 2015 Sep 28. J Biol Chem. 2015. PMID: 26416890 Free PMC article.
-
Substrate specificity of the ubiquitin and Ubl proteases.Cell Res. 2016 Apr;26(4):441-56. doi: 10.1038/cr.2016.38. Epub 2016 Mar 25. Cell Res. 2016. PMID: 27012468 Free PMC article. Review.
References
-
- Ciechanover A. Proteolysis: from the lysosome to ubiquitin and the proteasome. Nat Rev Mol Cell Biol. 2005;6:79–87. - PubMed
-
- Ciechanover A, Schwartz AL. Ubiquitin-mediated degradation of cellular proteins in health and disease. Hepatology. 2002;35:3–6. - PubMed
-
- Varshavsky A. The ubiquitin system. Trends Biochem Sci. 1997;22:383–387. - PubMed
-
- Wilkinson KD. Ubiquitination and deubiquitination: targeting of proteins for degradation by the proteasome. Semin Cell Dev Biol. 2000;11:141–148. - PubMed
-
- Goldberg AL. Protein degradation and protection against misfolded or damaged proteins. Nature. 2003;426:895–899. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
