

Temporal and spatial requirements of Smoothened in ventral midbrain neuronal development
- PMID: 23618354
- PMCID: PMC3680293
- DOI: 10.1186/1749-8104-8-8
Temporal and spatial requirements of Smoothened in ventral midbrain neuronal development
Abstract
Background: Several studies have indicated that Sonic hedgehog (Shh) regulates the expansion of dopaminergic (DA) progenitors and the subsequent generation of mature DA neurons. This prevailing view has been based primarily on in vitro culture results, and the exact in vivo function of Shh signaling in the patterning and neurogenesis of the ventral midbrain (vMB) remains unclear.
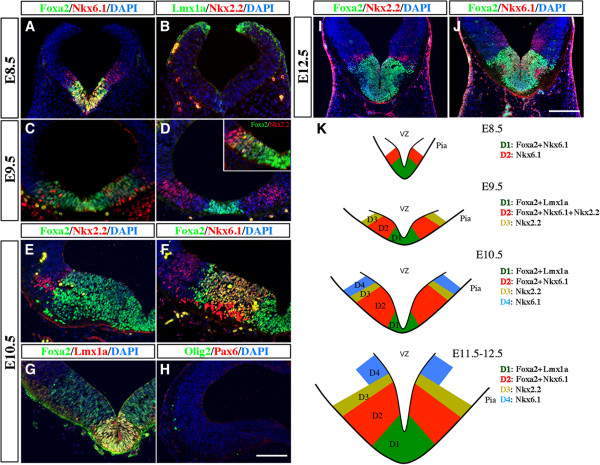
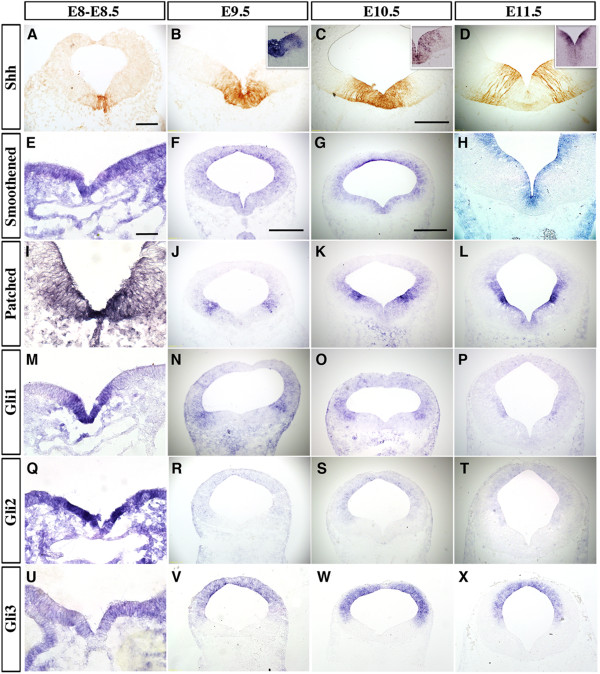
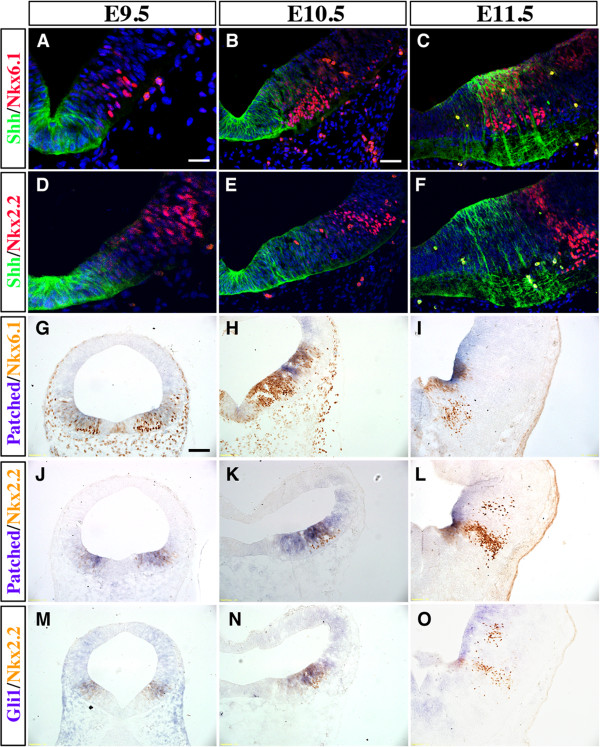
Methods: We characterized the transcriptional codes for the vMB progenitor domains, and correlated them with the expression patterns of Shh signaling effectors, including Shh, Smoothened, Patched, Gli1, Gli2 and Gli3.
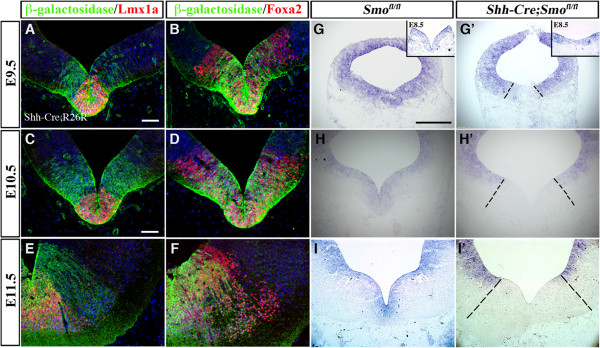
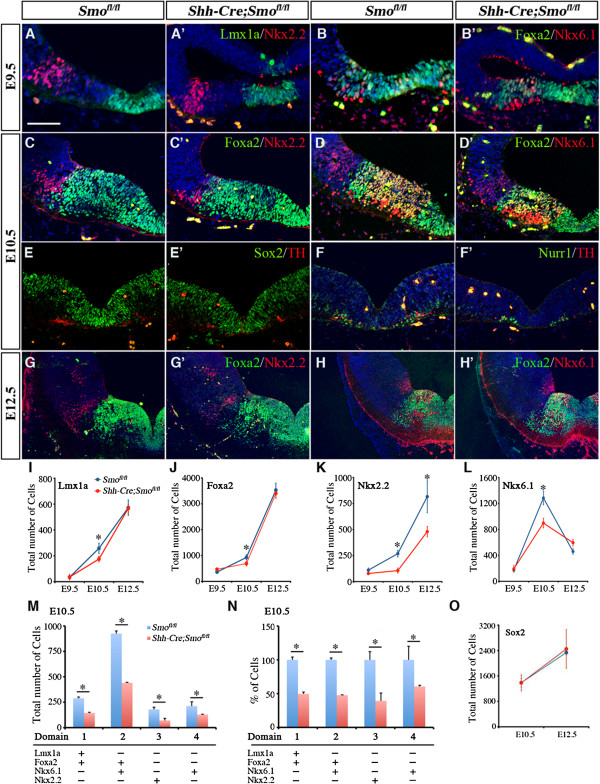
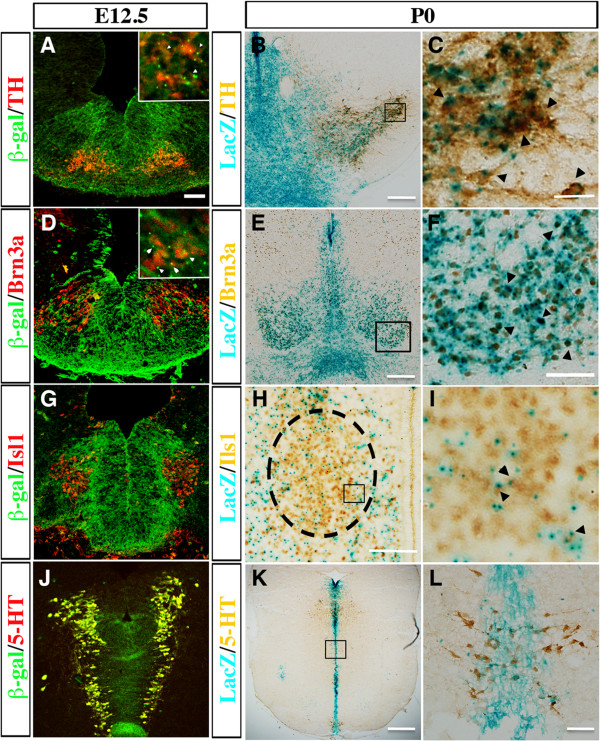
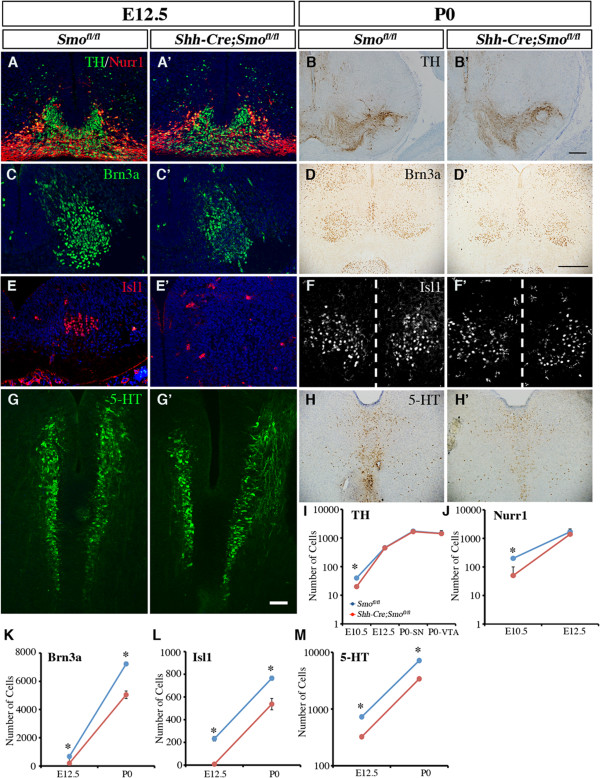
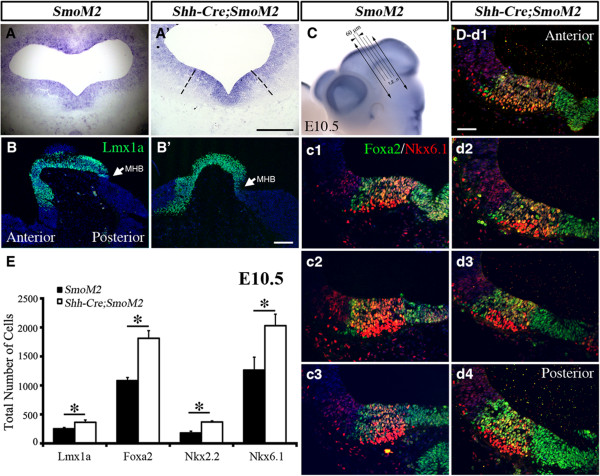
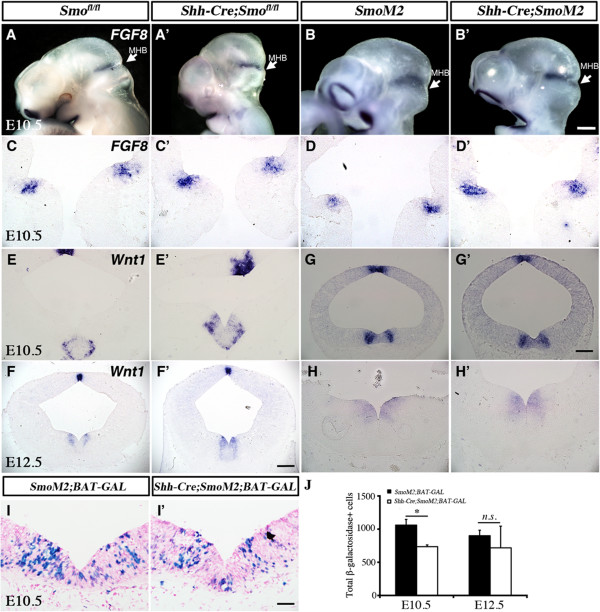
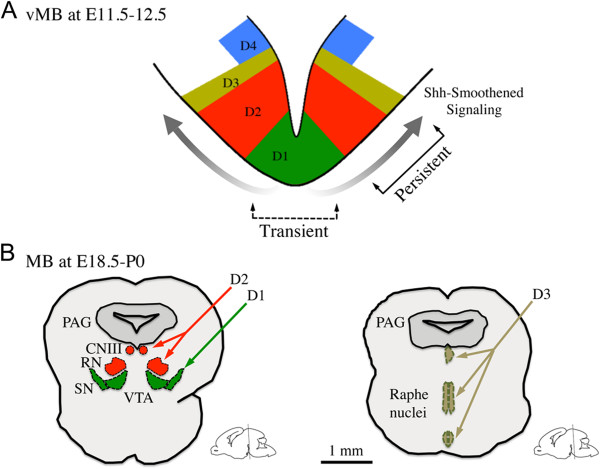
Results: While Shh and its downstream effectors showed robust expression in the neurogenic niche for DA progenitors at embryonic day (E)8 to E8.5, their expression shifted to the lateral domains from E9.5 to E12.5. Consistent with this dynamic change, conditional mutants with region-specific removal of the Shh receptor Smoothened in the vMB progenitors (Shh-Cre;Smo(fl/fl)) showed a transient reduction in DA progenitors and DA neurons at E10.5, but had more profound defects in neurons derived from the more lateral domains, including those in the red nucleus, oculomotor nucleus, and raphe nuclei. Conversely, constitutive activation of Smoothened signaling in vMB (Shh-Cre;SmoM2) showed transient expansion of the same progenitor population. To further characterize the nature of Shh-Smoothened signaling in vMB, we examined the BAT-GAL reporter and the expression of Wnt1 in vMB, and found that the antagonistic effects of Shh and Wnt signaling critically regulate the development of DA progenitors and DA neurons.
Conclusion: These results highlight previously unrecognized effects of Shh-Smoothened signaling in the region-specific neurogenesis within the vMB.
Figures
Similar articles
-
Primary cilia are critical for Sonic hedgehog-mediated dopaminergic neurogenesis in the embryonic midbrain.Dev Biol. 2016 Jan 1;409(1):55-71. doi: 10.1016/j.ydbio.2015.10.033. Epub 2015 Nov 2. Dev Biol. 2016. PMID: 26542012 Free PMC article.
-
Interactions of Wnt/beta-catenin signaling and sonic hedgehog regulate the neurogenesis of ventral midbrain dopamine neurons.J Neurosci. 2010 Jul 7;30(27):9280-91. doi: 10.1523/JNEUROSCI.0860-10.2010. J Neurosci. 2010. PMID: 20610763 Free PMC article.
-
N-cadherin regulates the proliferation and differentiation of ventral midbrain dopaminergic progenitors.Dev Neurobiol. 2013 Jul;73(7):518-29. doi: 10.1002/dneu.22077. Epub 2013 May 14. Dev Neurobiol. 2013. PMID: 23420609
-
Small-molecule modulators of the Sonic Hedgehog signaling pathway.Mol Biosyst. 2010 Jan;6(1):44-54. doi: 10.1039/b910196a. Epub 2009 Aug 27. Mol Biosyst. 2010. PMID: 20024066 Review.
-
The role of Smo-Shh/Gli signaling activation in the prevention of neurological and ageing disorders.Biogerontology. 2023 Aug;24(4):493-531. doi: 10.1007/s10522-023-10034-1. Epub 2023 Apr 25. Biogerontology. 2023. PMID: 37097427 Review.
Cited by
-
Multi-omic analysis of selectively vulnerable motor neuron subtypes implicates altered lipid metabolism in ALS.Nat Neurosci. 2021 Dec;24(12):1673-1685. doi: 10.1038/s41593-021-00944-z. Epub 2021 Nov 15. Nat Neurosci. 2021. PMID: 34782793 Free PMC article.
-
N-cadherin-based adherens junction regulates the maintenance, proliferation, and differentiation of neural progenitor cells during development.Cell Adh Migr. 2015;9(3):183-92. doi: 10.1080/19336918.2015.1005466. Epub 2015 Apr 14. Cell Adh Migr. 2015. PMID: 25869655 Free PMC article. Review.
-
Constitutive and conditional gene knockout mice for the study of intervertebral disc degeneration: Current status, decision considerations, and future possibilities.JOR Spine. 2023 Jan 7;6(1):e1242. doi: 10.1002/jsp2.1242. eCollection 2023 Mar. JOR Spine. 2023. PMID: 36994464 Free PMC article. Review.
-
Primary cilia are critical for Sonic hedgehog-mediated dopaminergic neurogenesis in the embryonic midbrain.Dev Biol. 2016 Jan 1;409(1):55-71. doi: 10.1016/j.ydbio.2015.10.033. Epub 2015 Nov 2. Dev Biol. 2016. PMID: 26542012 Free PMC article.
-
Definition of a critical spatiotemporal window within which primary cilia control midbrain dopaminergic neurogenesis.Neurogenesis (Austin). 2016 Oct 20;3(1):e1248206. doi: 10.1080/23262133.2016.1248206. eCollection 2016. Neurogenesis (Austin). 2016. PMID: 28090543 Free PMC article.
References
-
- Chamberlain CE, Jeong J, Guo C, Allen BL, McMahon AP. Notochord-derived Shh concentrates in close association with the apically positioned basal body in neural target cells and forms a dynamic gradient during neural patterning. Development. 2008;135:1097–1106. doi: 10.1242/dev.013086. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
