

Review
doi: 10.1038/nri3423.
Beyond oxidative stress: an immunologist's guide to reactive oxygen species
Affiliations
- PMID: 23618831
- PMCID: PMC4250048
- DOI: 10.1038/nri3423
Item in Clipboard
Review
Beyond oxidative stress: an immunologist's guide to reactive oxygen species
Nat Rev Immunol.
2013 May.
Abstract
Reactive oxygen species (ROS) react preferentially with certain atoms to modulate functions ranging from cell homeostasis to cell death. Molecular actions include both inhibition and activation of proteins, mutagenesis of DNA and activation of gene transcription. Cellular actions include promotion or suppression of inflammation, immunity and carcinogenesis. ROS help the host to compete against microorganisms and are also involved in intermicrobial competition. ROS chemistry and their pleiotropy make them difficult to localize, to quantify and to manipulate - challenges we must overcome to translate ROS biology into medical advances.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
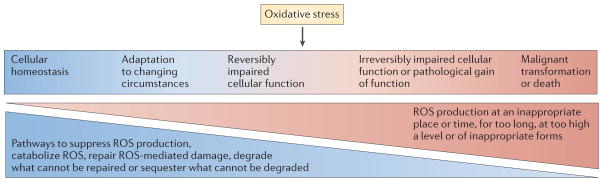
Restriction of reactive oxygen species (ROS) production to appropriate subcellular locations, times, levels, molecular species and for appropriate durations allows ROS to contribute to homeostasis and physiological cell activation. For example, brief pulses of H2O2 production at the plasma membrane or at the endosomal membrane mediate signalling in response to the engagement of receptors with cytokines, microbial products or antigens (left-hand side). When ROS production escapes these restrictions — for example, when there are high levels or sustained production of hydroxyl radicals — macromolecules are damaged (‘oxidative stress’). ROS-mediated damage can often be reversed by repair, replacement, degradation or sequestration of the damaged macromolecules (middle). However, damage that exceeds the capacity of the cell for these responses can lead to cell death (right-hand side). When damage to DNA results in mutagenesis without irreparable double-strand breakage, and when damage to other macromolecules is repaired, the consequence can be malignant transformation rather than death of the cell (right-hand side).
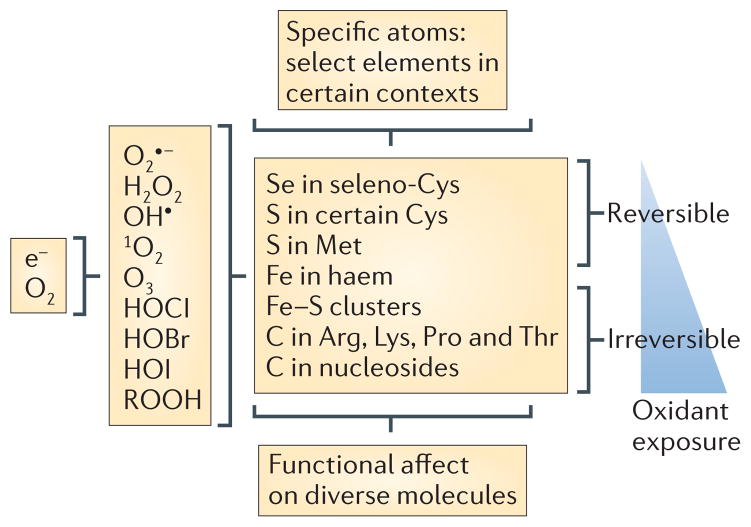
During the reduction of oxygen to water, sequential one-electron reductions can produce reactive oxygen intermediates (ROIs) — superoxide, hydrogen peroxide and hydroxyl radicals — along with singlet oxygen and ozone. ROIs comprise a subset of reactive oxygen species (ROS). Additional ROS are the hypochlorous (HOCl), hypobromous (HOBr) and hypoiodous acids (HOI) that arise when peroxidases catalyse the oxidation of halides by H2O2, as well as important products of the reaction of ROS with other molecules that retain strong oxidizing potential, such as lipid peroxides (included as ROOH in the figure). ROS at low levels tend to react reversibly with a limited number of atoms — for example, selenium or sulphur atoms in a subset of cysteine and methionine residues — conferring atomic specificity in reactions involving diverse macromolecules. At higher levels ROS are likely to react irreversibly with certain iron and carbon atoms. e−, electron.
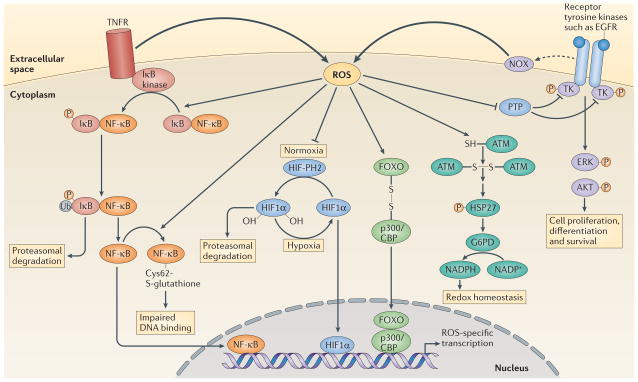
Activation of tumour necrosis factor receptor (TNFR) triggers reactive oxygen species (ROS) production, which enhances the phosphorylation (P) of inhibitor of NF-κB (IκB), probably through the oxidative inhibition of a phosphatase. This leads to the ubiquitylation (Ub) of IκB and its subsequent degradation by the proteasome. Nuclear factor-κB (NF-κB) is then released and translocates to the nucleus to initiate transcription. ROS production can also trigger oxidative glutathionylation of NF-κB at its redox sensitive cysteine, which reduces its DNA binding affinity. Under normoxia, prolyl hydroxylases (PHs) hydroxylate hypoxia-inducible factor 1α (HIF11α), which allows it to be recognized by the E3 ligase von Hippel–Lindau tumour-suppressor protein (VHL) and promotes its degradation by the proteasome. Under hypoxia there is increased production of ROS (FIG. 4), which inhibits prolyl hydroxylases, leading to the accumulation of HIF11α. HIF11α then translocates to the nucleus to mediate gene transcription. ROS production by NADPH oxidases (NOXs) following receptor activation by specific ligands, for example, epidermal growth factor receptor (EGFR), inhibits protein tyrosine phosphatases (PTPs), which promotes the phosphorylation of tyrosine kinases (TKs) and the subsequent signal transduction. By contrast, ataxia-telangiectasia mutated (ATM) kinase is activated directly by ROS, through disulphide bond-mediated homodimerization, which leads to the phosphorylation of heat shock protein 27 (HSP27) and the subsequent activation of glucose-6-phosphate-dehydrogenase (G6PD). The resulting increase in NADPH levels contributes to the maintenance of cellular redox homeostasis. ROS-mediated disulphide bonding can also lead to heterodimerization, as seen between forkhead box O (FOXO) transcription factors and p300/CBP acetyltransferase, which leads to the acetylation of FOXO proteins and the activation of specific gene transcription. ERK, extracellular signal-regulated kinase.
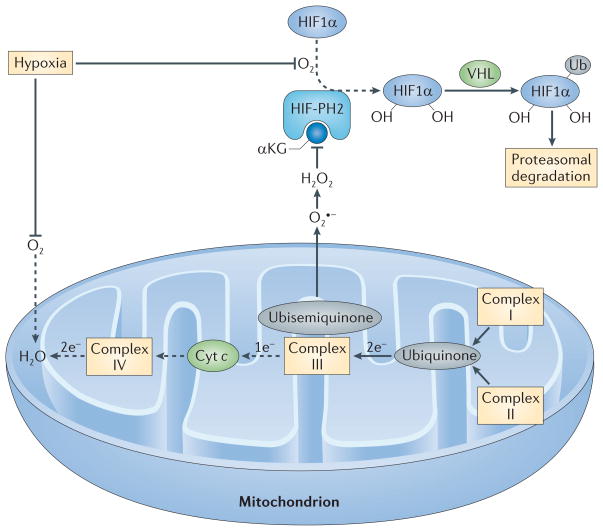
In the presence of O2 and its cofactor α-ketoglutarate (αKG), HIF-prolyl hydroxylase 2 (HIF-PH2) hydroxylates two proline residues in hypoxia-inducible factor 1α (HIF1α). Hydroxylated HIF1α is then ubiquitylated (Ub) by the E3 ligase von Hippel–Lindau tumour-suppressor protein (VHL) and is subsequently degraded by the proteasome. Because O2 is a substrate for HIF-PH2, hypoxia limits HIF-PH2 activity. This limitation is enhanced by the negative effect of hypoxia-driven mitochondrial reactive oxygen species (ROS) on HIF-PH2 function. Complex III in the mitochondrial electron transport chain (METC) receives two electrons from ubiquinone but can only transfer one electron at a time to cytochrome c (Cyt c). Complex III therefore transfers one electron to a quasi-stable ubisemiquinone radical. If this radical accumulates, O2 that is dissolved in the mitochondrial membrane can capture the electron before cytochrome c accepts it, generating superoxide (O2•−). If cellular O2 is low, complex IV at the end of the METC is slow to transfer pairs of electrons to O2 (making water), and the METC is blocked between complex III and IV, which favours the electron leak described above. The newly formed superoxide dismutates to H2O2. H2O2 can inhibit HIF-PH2 and oxidatively decarboxylate its cofactor, αKG. The resulting decrease in the hydroxylation of HIF1α and its subsequent proteasomal degradation supports HIF1α accumulation. Dashed arrows indicate the reactions that are slowed in hypoxia.
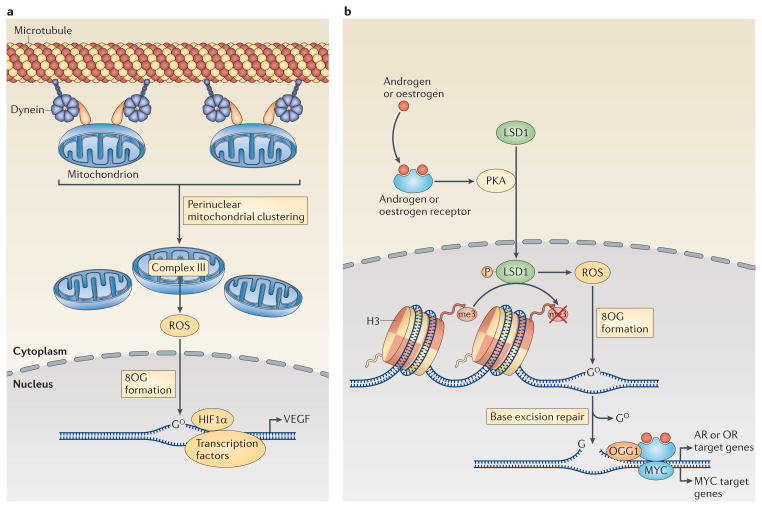
a | The induction of the transcription of the gene encoding vascular endothelial growth factor (VEGF) by hypoxia-inducible factor 1α (HIF1α) is enhanced through the dynein-mediated, perinuclear localization of mitochondria. Mitochondria-derived reactive oxygen species (ROS) diffuse into the nucleus, where they promote the oxidation of guanine nucleotides, forming 8-oxoguanine (8OG). b | Transcriptional regulation downstream of the activation of androgen receptors (ARs) or oestrogen receptors (ORs) and other nuclear receptors also involves DNA modifications by ROS. Engagement of ORs or ARs promotes the phosphorylation (P) of lysine-specific histone demethylase 1A (LSD1) by cAMP-dependent protein kinase (PKA). Active LSD1 not only demethylates histone 3 (H3) but also produces ROS, which then promote the formation of 8OG in the DNA. The altered DNA bases recruit base excision repair machinery, and the DNA breaks that are generated by 8OG DNA glycosylase 1 (OGG1) enable the activation of transcription by AR, OR and possibly also MYC.
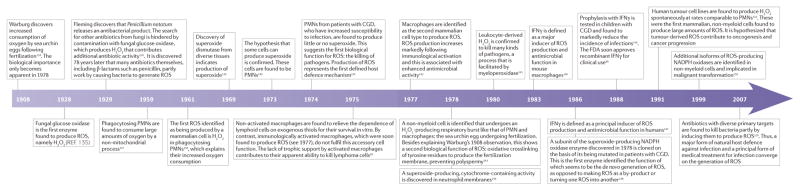
A sampling of milestones in ROS biology* CGD, chronic granulomatous disease; FDA, US Food and Drug Administration; IFNγ, interferon-γ; PMN, polymorphonuclear leukocyte; ROS, reactive oxygen species. *This Timeline is an incomplete history, with only limited citations.
References
-
- Nathan C, Ding A. Snapshot: reactive oxygen intermediates (ROI) Cell. 2010;140:951–951.e2. - PubMed
-
- Finkel T. From sulfenylation to sulfhydration: what a thiolate needs to tolerate. Sci Signal. 2012;5:pe10. - PubMed
-
- Paul BD, Snyder SH. H2S signalling through protein sulfhydration and beyond. Nature Rev Mol Cell Biol. 2012;13:499–507. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
