

TnaA, an SP-RING protein, interacts with Osa, a subunit of the chromatin remodeling complex BRAHMA and with the SUMOylation pathway in Drosophila melanogaster
- PMID: 23620817
- PMCID: PMC3631182
- DOI: 10.1371/journal.pone.0062251
TnaA, an SP-RING protein, interacts with Osa, a subunit of the chromatin remodeling complex BRAHMA and with the SUMOylation pathway in Drosophila melanogaster
Abstract
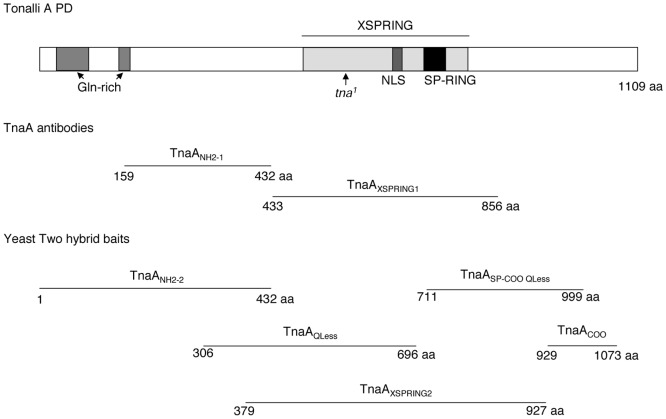
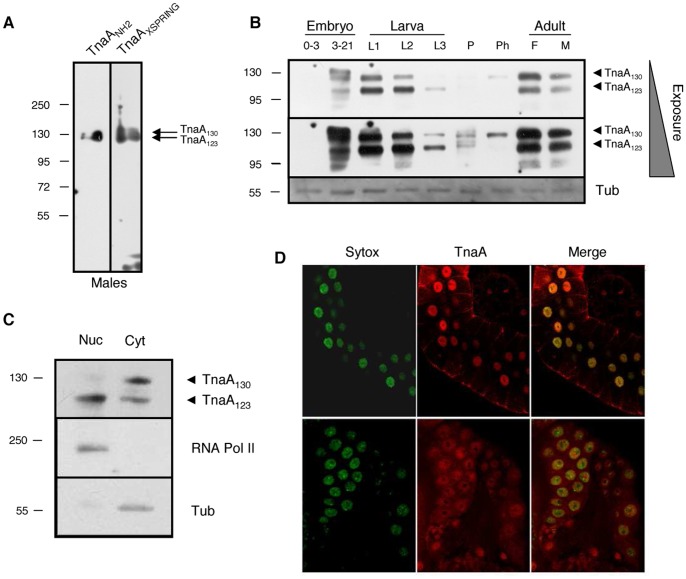
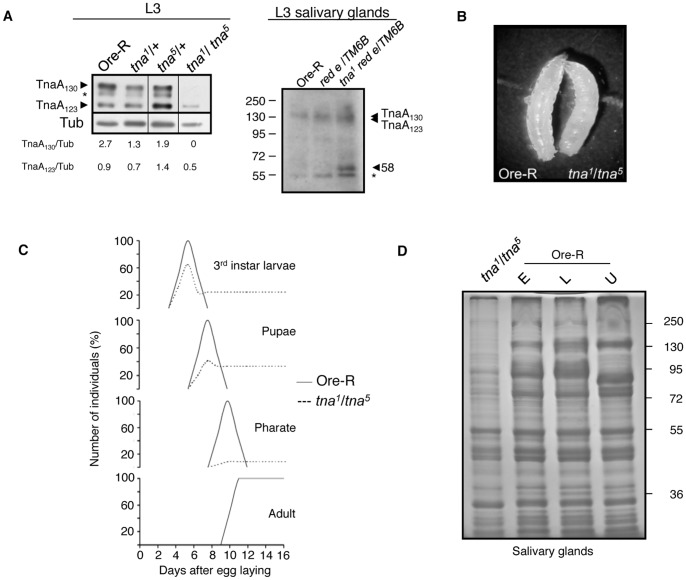
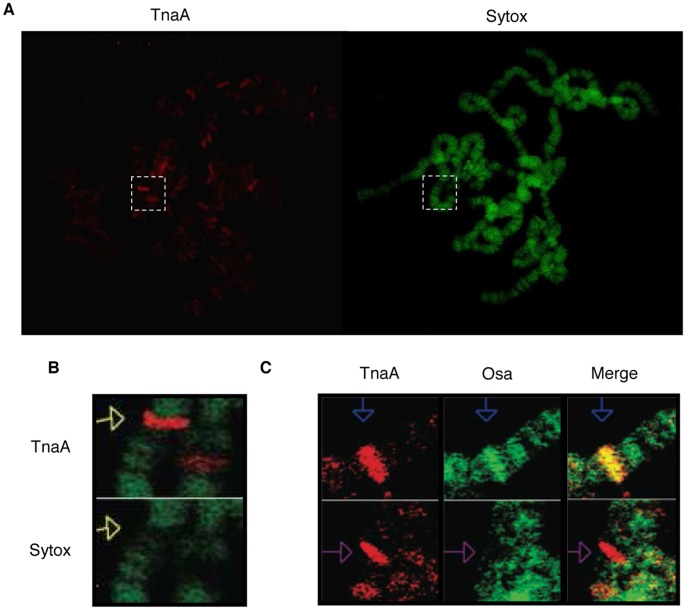
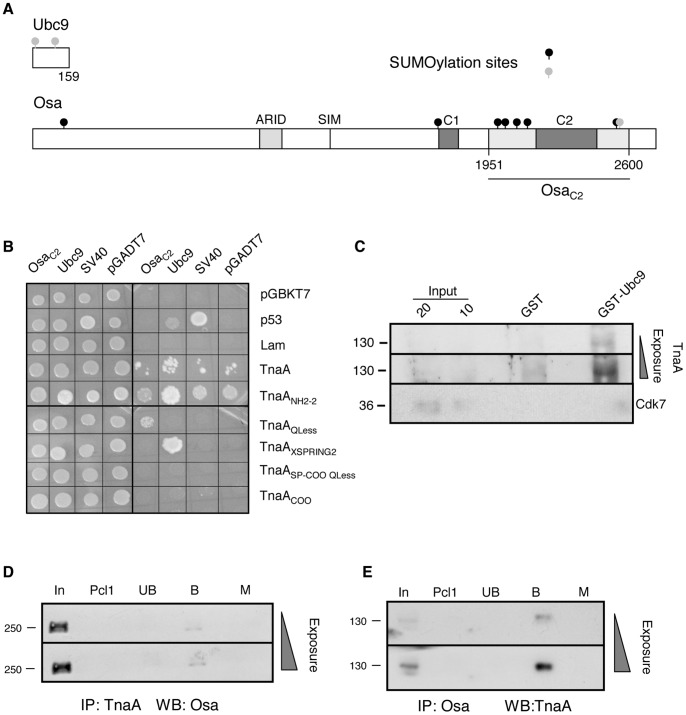
Tonalli A (TnaA) is a Drosophila melanogaster protein with an XSPRING domain. The XSPRING domain harbors an SP-RING zinc-finger, which is characteristic of proteins with SUMO E3 ligase activity. TnaA is required for homeotic gene expression and is presumably involved in the SUMOylation pathway. Here we analyzed some aspects of the TnaA location in embryo and larval stages and its genetic and biochemical interaction with SUMOylation pathway proteins. We describe that there are at least two TnaA proteins (TnaA130 and TnaA123) differentially expressed throughout development. We show that TnaA is chromatin-associated at discrete sites on polytene salivary gland chromosomes of third instar larvae and that tna mutant individuals do not survive to adulthood, with most dying as third instar larvae or pupae. The tna mutants that ultimately die as third instar larvae have an extended life span of at least 4 to 15 days as other SUMOylation pathway mutants. We show that TnaA physically interacts with the SUMO E2 conjugating enzyme Ubc9, and with the BRM complex subunit Osa. Furthermore, we show that tna and osa interact genetically with SUMOylation pathway components and individuals carrying mutations for these genes show a phenotype that can be the consequence of misexpression of developmental-related genes.
Conflict of interest statement
Figures

Similar articles
-
The Drosophila trithorax group gene tonalli (tna) interacts genetically with the Brahma remodeling complex and encodes an SP-RING finger protein.Development. 2003 Jan;130(2):343-54. doi: 10.1242/dev.00222. Development. 2003. PMID: 12466201
-
The role of the trithorax group TnaA isoforms in Hox gene expression, and in Drosophila late development.PLoS One. 2018 Oct 29;13(10):e0206587. doi: 10.1371/journal.pone.0206587. eCollection 2018. PLoS One. 2018. PMID: 30372466 Free PMC article.
-
The Drosophila SNR1 (SNF5/INI1) subunit directs essential developmental functions of the Brahma chromatin remodeling complex.Mol Cell Biol. 2003 Jan;23(1):289-305. doi: 10.1128/MCB.23.1.289-305.2003. Mol Cell Biol. 2003. PMID: 12482982 Free PMC article.
-
Differential targeting of two distinct SWI/SNF-related Drosophila chromatin-remodeling complexes.Mol Cell Biol. 2004 Apr;24(8):3077-88. doi: 10.1128/MCB.24.8.3077-3088.2004. Mol Cell Biol. 2004. PMID: 15060132 Free PMC article.
-
SUMO in Drosophila Development.Adv Exp Med Biol. 2017;963:249-257. doi: 10.1007/978-3-319-50044-7_15. Adv Exp Med Biol. 2017. PMID: 28197917 Review.
Cited by
-
SUMOylation of Warts kinase promotes neural stem cell reactivation.Nat Commun. 2024 Oct 17;15(1):8557. doi: 10.1038/s41467-024-52569-y. Nat Commun. 2024. PMID: 39419973 Free PMC article.
-
TnaA, a trithorax group protein, modulates wingless expression in different regions of the Drosophila wing imaginal disc.Sci Rep. 2023 Sep 13;13(1):15162. doi: 10.1038/s41598-023-42169-z. Sci Rep. 2023. PMID: 37704704 Free PMC article.
-
A comprehensive platform for the analysis of ubiquitin-like protein modifications using in vivo biotinylation.Sci Rep. 2017 Jan 18;7:40756. doi: 10.1038/srep40756. Sci Rep. 2017. PMID: 28098257 Free PMC article.
-
Transcriptional regulation by trithorax-group proteins.Cold Spring Harb Perspect Biol. 2014 Oct 1;6(10):a019349. doi: 10.1101/cshperspect.a019349. Cold Spring Harb Perspect Biol. 2014. PMID: 25274705 Free PMC article. Review.
-
SUMOylation in development and neurodegeneration.Development. 2020 Mar 18;147(6):dev175703. doi: 10.1242/dev.175703. Development. 2020. PMID: 32188601 Free PMC article. Review.
References
-
- Geiss-Friedlander R, Melchior F (2007) Concepts in SUMOylation: a decade on. Nat Rev Mol Cell Biol 10: 947–956. - PubMed
-
- Rodríguez MS, Dargemont C, Hay RT (2001) SUMO-1 conjugation in vivo requires both a consensus modification motif and nuclear targeting. J Biol Chem 276: 12654–12659. - PubMed
-
- Gutiérrez L, Zurita M, Kennison JA, Vázquez M (2003) The Drosophila trithorax group gene tonalli (tna) interacts genetically with the Brahma remodeling complex and encodes an SP-RING finger protein. Development 130: 343–354. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
