

Genomic selection of purebred animals for crossbred performance in the presence of dominant gene action
- PMID: 23621868
- PMCID: PMC3673865
- DOI: 10.1186/1297-9686-45-11
Genomic selection of purebred animals for crossbred performance in the presence of dominant gene action
Abstract
Background: Genomic selection is an appealing method to select purebreds for crossbred performance. In the case of crossbred records, single nucleotide polymorphism (SNP) effects can be estimated using an additive model or a breed-specific allele model. In most studies, additive gene action is assumed. However, dominance is the likely genetic basis of heterosis. Advantages of incorporating dominance in genomic selection were investigated in a two-way crossbreeding program for a trait with different magnitudes of dominance. Training was carried out only once in the simulation.
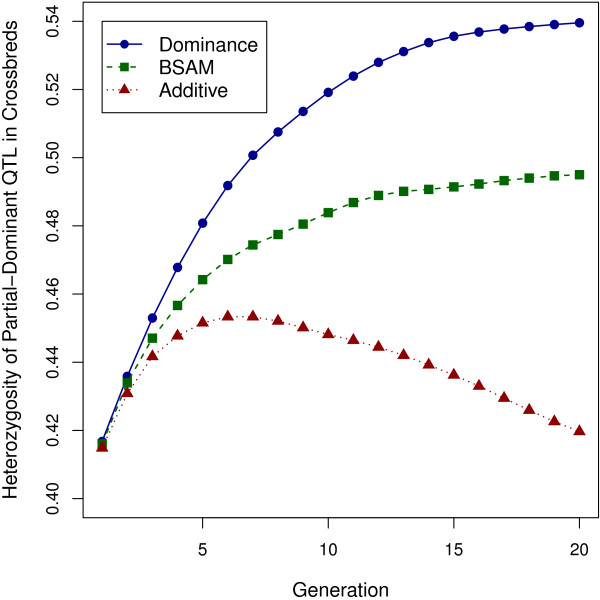
Results: When the dominance variance and heterosis were large and overdominance was present, a dominance model including both additive and dominance SNP effects gave substantially greater cumulative response to selection than the additive model. Extra response was the result of an increase in heterosis but at a cost of reduced purebred performance. When the dominance variance and heterosis were realistic but with overdominance, the advantage of the dominance model decreased but was still significant. When overdominance was absent, the dominance model was slightly favored over the additive model, but the difference in response between the models increased as the number of quantitative trait loci increased. This reveals the importance of exploiting dominance even in the absence of overdominance. When there was no dominance, response to selection for the dominance model was as high as for the additive model, indicating robustness of the dominance model. The breed-specific allele model was inferior to the dominance model in all cases and to the additive model except when the dominance variance and heterosis were large and with overdominance. However, the advantage of the dominance model over the breed-specific allele model may decrease as differences in linkage disequilibrium between the breeds increase. Retraining is expected to reduce the advantage of the dominance model over the alternatives, because in general, the advantage becomes important only after five or six generations post-training.
Conclusion: Under dominance and without retraining, genomic selection based on the dominance model is superior to the additive model and the breed-specific allele model to maximize crossbred performance through purebred selection.
Figures





References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
