

Four-dimensional imaging of E. coli nucleoid organization and dynamics in living cells
- PMID: 23623305
- PMCID: PMC3670778
- DOI: 10.1016/j.cell.2013.04.006
Four-dimensional imaging of E. coli nucleoid organization and dynamics in living cells
Abstract
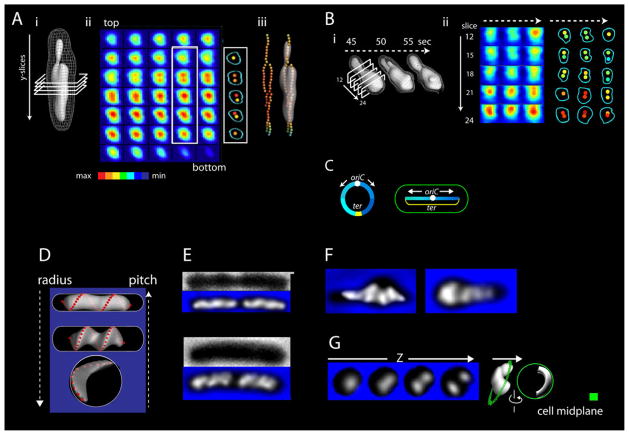
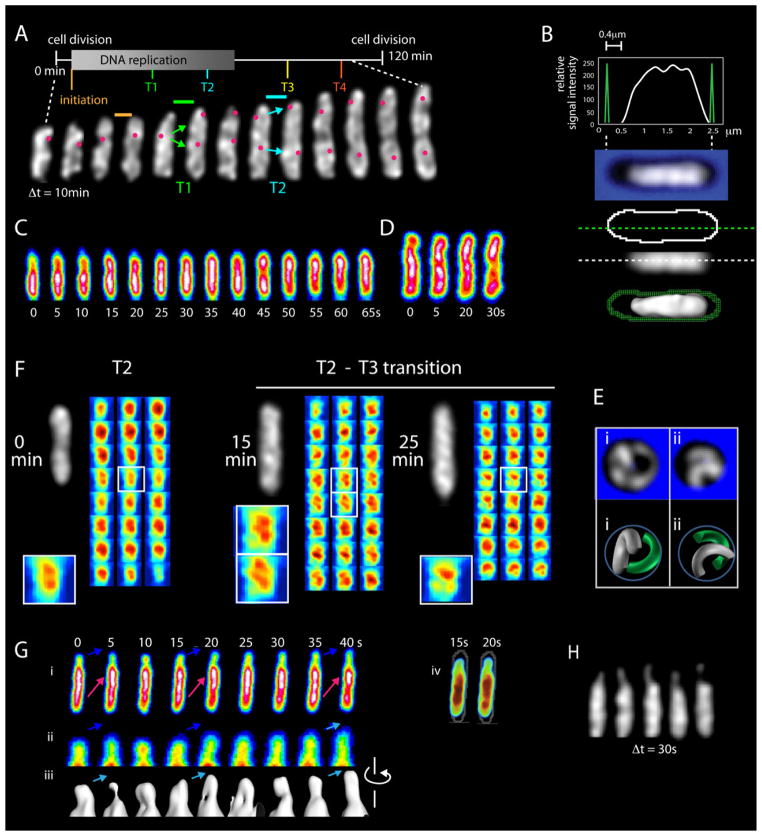
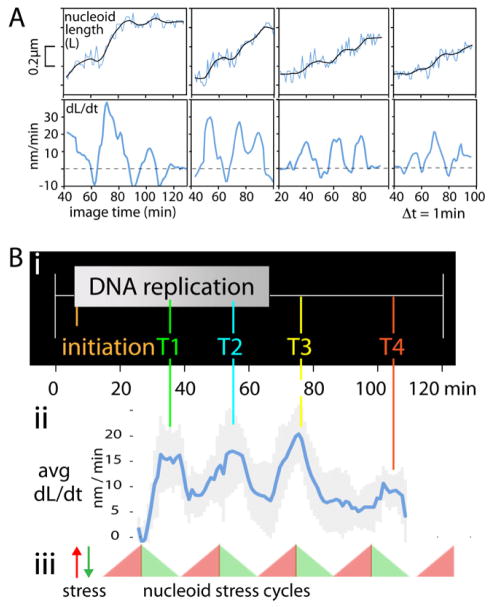
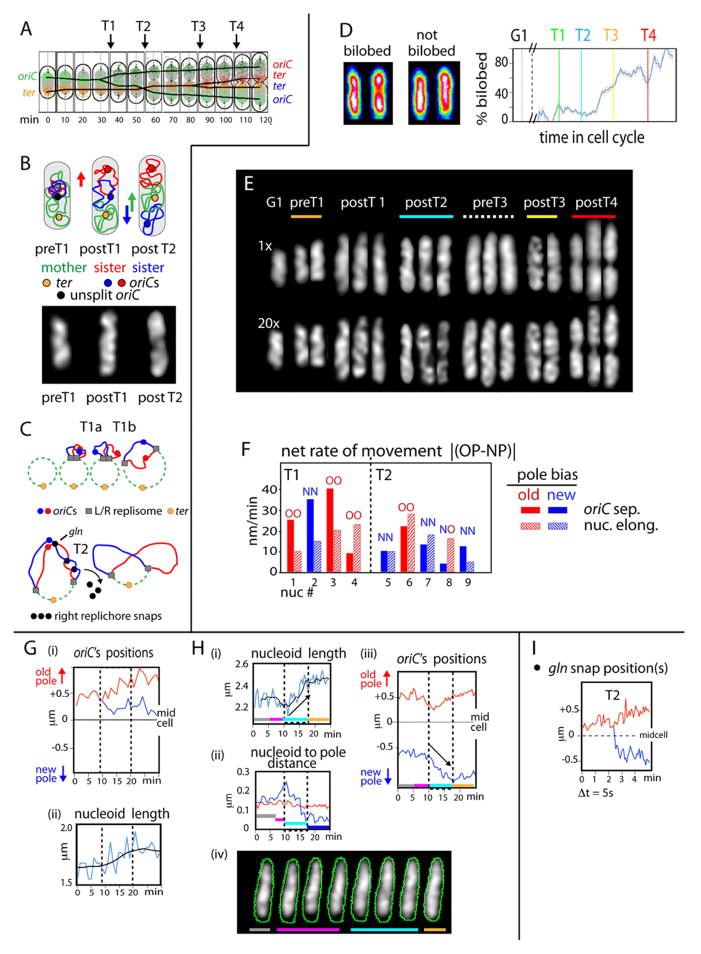
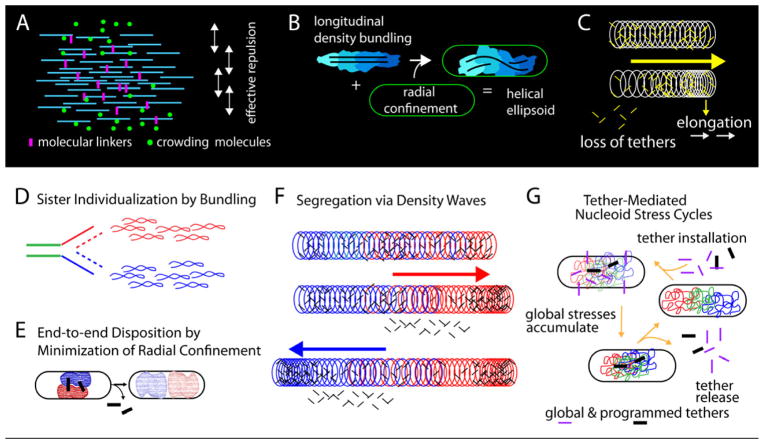
Visualization of living E. coli nucleoids, defined by HupA-mCherry, reveals a discrete, dynamic helical ellipsoid. Three basic features emerge. (1) Nucleoid density coalesces into longitudinal bundles, giving a stiff, low-DNA-density ellipsoid. (2) This ellipsoid is radially confined within the cell cylinder. Radial confinement gives helical shape and directs global nucleoid dynamics, including sister segregation. (3) Longitudinal density waves flux back and forth along the nucleoid, with 5%-10% of density shifting within 5 s, enhancing internal nucleoid mobility. Furthermore, sisters separate end-to-end in sequential discontinuous pulses, each elongating the nucleoid by 5%-15%. Pulses occur at 20 min intervals, at defined cell-cycle times. This progression includes sequential installation and release of programmed tethers, implying cyclic accumulation and relief of intranucleoid mechanical stress. These effects could comprise a chromosome-based cell-cycle engine. Overall, the presented results suggest a general conceptual framework for bacterial nucleoid morphogenesis and dynamics.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures

Comment in
-
A close look at wiggly chromosomes.Dev Cell. 2013 May 28;25(4):330-2. doi: 10.1016/j.devcel.2013.05.005. Dev Cell. 2013. PMID: 23725761 Free PMC article.
Similar articles
-
Escherichia coli sister chromosome separation includes an abrupt global transition with concomitant release of late-splitting intersister snaps.Proc Natl Acad Sci U S A. 2011 Feb 15;108(7):2765-70. doi: 10.1073/pnas.1019593108. Epub 2011 Jan 31. Proc Natl Acad Sci U S A. 2011. PMID: 21282646 Free PMC article.
-
The bacterial nucleoid: nature, dynamics and sister segregation.Curr Opin Microbiol. 2014 Dec;22:127-37. doi: 10.1016/j.mib.2014.10.001. Curr Opin Microbiol. 2014. PMID: 25460806 Free PMC article. Review.
-
The Origin of Chromosomal Replication Is Asymmetrically Positioned on the Mycobacterial Nucleoid, and the Timing of Its Firing Depends on HupB.J Bacteriol. 2018 Apr 24;200(10):e00044-18. doi: 10.1128/JB.00044-18. Print 2018 May 15. J Bacteriol. 2018. PMID: 29531181 Free PMC article.
-
Cell Boundary Confinement Sets the Size and Position of the E. coli Chromosome.Curr Biol. 2019 Jul 8;29(13):2131-2144.e4. doi: 10.1016/j.cub.2019.05.015. Epub 2019 May 30. Curr Biol. 2019. PMID: 31155353 Free PMC article.
-
Nucleoid-mediated positioning and transport in bacteria.Curr Genet. 2020 Apr;66(2):279-291. doi: 10.1007/s00294-019-01041-2. Epub 2019 Nov 5. Curr Genet. 2020. PMID: 31691024 Review.
Cited by
-
Emergent perversions in the buckling of heterogeneous elastic strips.Proc Natl Acad Sci U S A. 2016 Jun 28;113(26):7100-5. doi: 10.1073/pnas.1605621113. Epub 2016 Jun 14. Proc Natl Acad Sci U S A. 2016. PMID: 27303040 Free PMC article.
-
Regional Control of Chromosome Segregation in Pseudomonas aeruginosa.PLoS Genet. 2016 Nov 7;12(11):e1006428. doi: 10.1371/journal.pgen.1006428. eCollection 2016 Nov. PLoS Genet. 2016. PMID: 27820816 Free PMC article.
-
The E. coli MinCDE system in the regulation of protein patterns and gradients.Cell Mol Life Sci. 2019 Nov;76(21):4245-4273. doi: 10.1007/s00018-019-03218-x. Epub 2019 Jul 17. Cell Mol Life Sci. 2019. PMID: 31317204 Free PMC article. Review.
-
Supercoiling and looping promote DNA base accessibility and coordination among distant sites.Nat Commun. 2021 Sep 28;12(1):5683. doi: 10.1038/s41467-021-25936-2. Nat Commun. 2021. PMID: 34584096 Free PMC article.
-
Mechanisms for Chromosome Segregation in Bacteria.Front Microbiol. 2021 Jun 16;12:685687. doi: 10.3389/fmicb.2021.685687. eCollection 2021. Front Microbiol. 2021. PMID: 34220773 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
