

dusky-like is required to maintain the integrity and planar cell polarity of hairs during the development of the Drosophila wing
- PMID: 23623898
- PMCID: PMC3686509
- DOI: 10.1016/j.ydbio.2013.04.012
dusky-like is required to maintain the integrity and planar cell polarity of hairs during the development of the Drosophila wing
Abstract
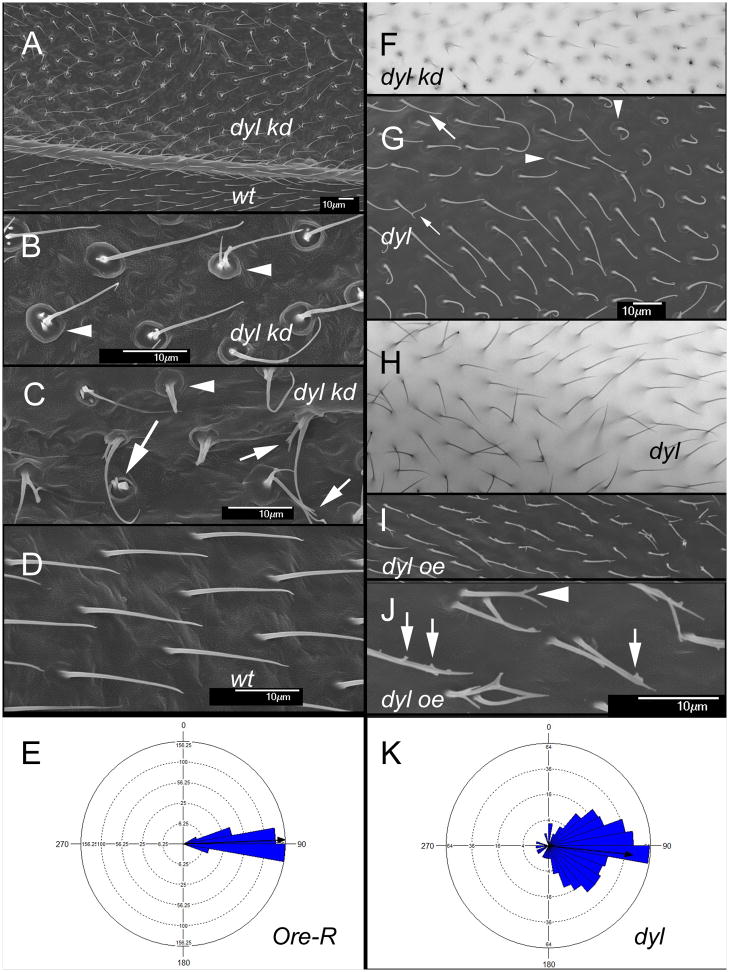
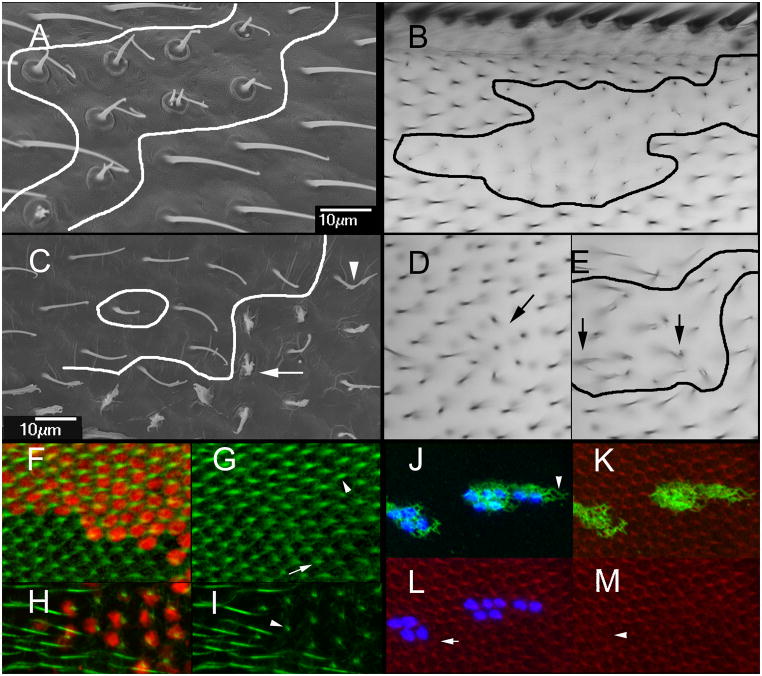
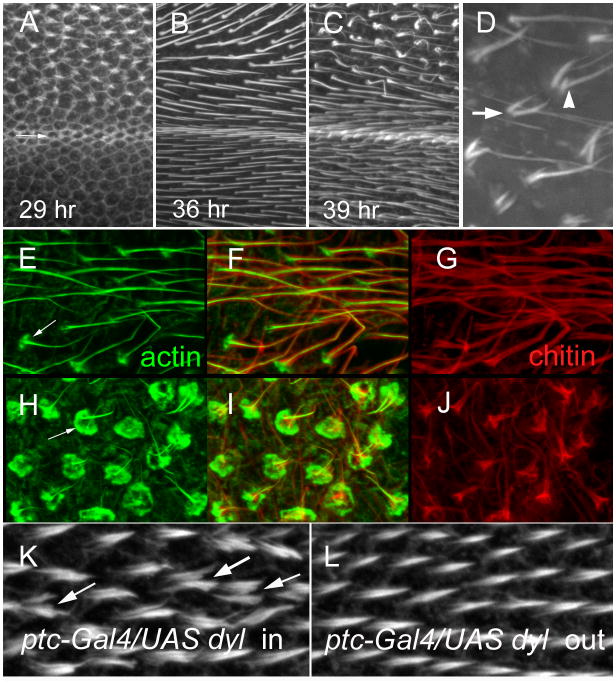
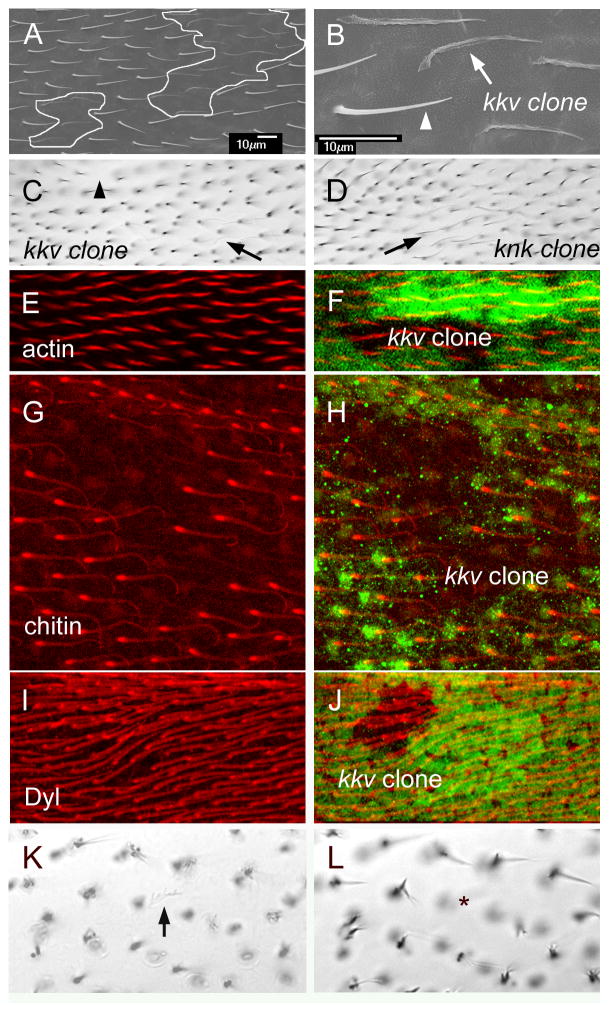
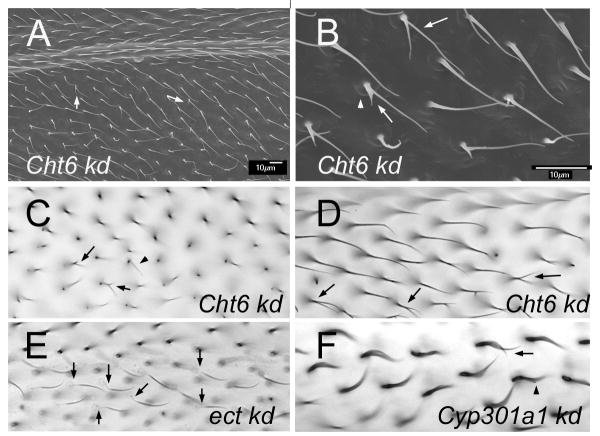
The cuticular hairs and sensory bristles that decorate the adult Drosophila epidermis and the denticles found on the embryo have been used in studies on planar cell polarity and as models for the cytoskeletal mediated morphogenesis of cellular extensions. ZP domain proteins have recently been found to be important for the morphogenesis of both denticles and bristles. Here we show that the ZP domain protein Dusky-like is a key player in hair morphogenesis. As is the case in bristles, in hairs dyl mutants display a dramatic phenotype that is the consequence of a failure to maintain the integrity of the extension after outgrowth. Hairs lacking dyl function are split, thinned, multipled and often very short. dyl is required for normal chitin deposition in hairs, but chitin is not required for the normal accumulation of Dyl, hence dyl acts upstream of chitin. A lack of chitin however, does not mimic the dyl hair phenotype, thus Dyl must have other targets in hair morphogenesis. One of these appears to be the actin cytoskeleton. Interestingly, dyl mutants also display a unique planar cell polarity phenotype that is distinct from that seen with mutations in the frizzled/starry night or dachsous/fat pathway genes. Rab11 was previously found to be essential for Dyl plasma membrane localization in bristles. Here we found that the expression of a dominant negative Rab11 can mimic the dyl hair morphology phenotype consistent with Rab11 also being required for Dyl function in hairs. We carried out a small directed screen to identify genes that might function with dyl and identified Chitinase 6 (Cht6) as a strong candidate, as knocking down Cht6 function led to weak versions of all of the dyl hair phenotypes.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures


References
-
- Adler PN. Planar signaling and morphogenesis in Drosophila. Dev Cell. 2002;2:525–535. - PubMed
-
- Adler PN, Charlton J, Liu J. Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype by altering frizzled signaling. Development. 1998;125:959–968. - PubMed
-
- Adler PN, Zhu C, Stone D. Inturned Localizes to the Proximal Side of Wing Cells under the Instruction of Upstream Planar Polarity Proteins. Curr Biol. 2004;14:2046–2051. - PubMed
-
- Amonlirdviman K, Khare NA, Tree DR, Chen WS, Axelrod JD, Tomlin CJ. Mathematical modeling of planar cell polarity to understand domineering nonautonomy. Science. 2005;307:423–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
