

Distribution of cp32 prophages among Lyme disease-causing spirochetes and natural diversity of their lipoprotein-encoding erp loci
- PMID: 23624478
- PMCID: PMC3697573
- DOI: 10.1128/AEM.00817-13
Distribution of cp32 prophages among Lyme disease-causing spirochetes and natural diversity of their lipoprotein-encoding erp loci
Abstract
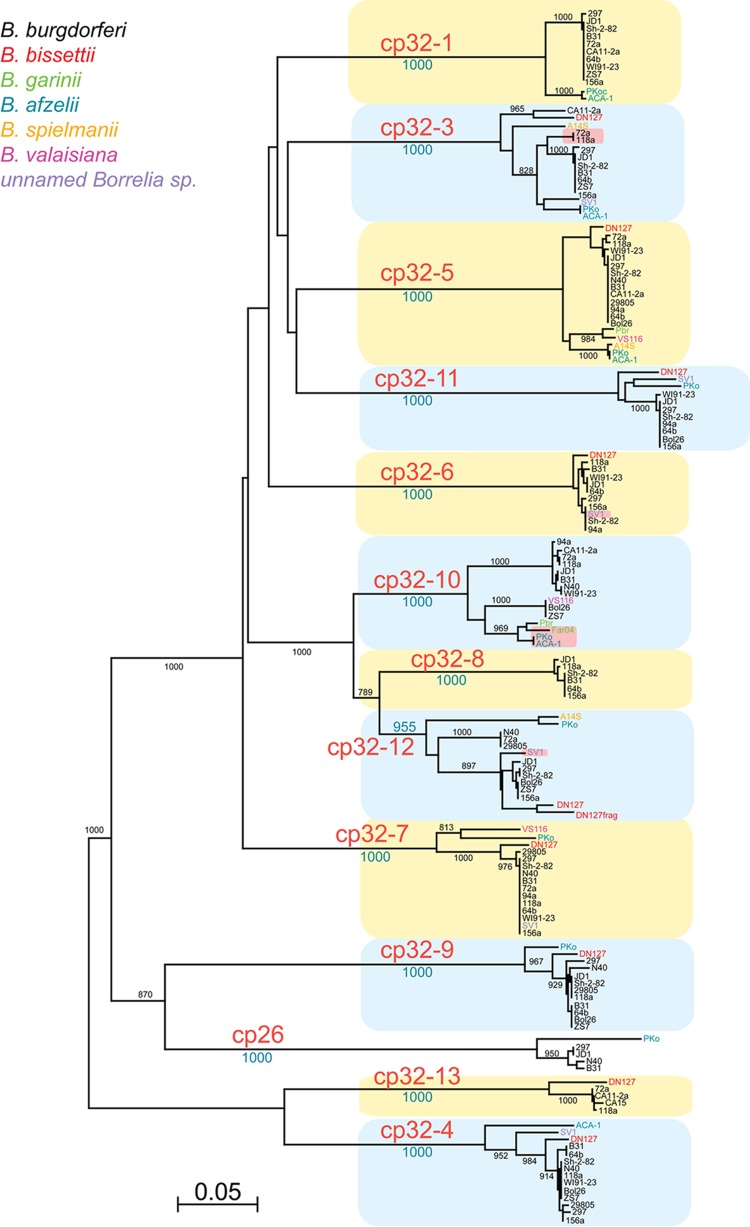
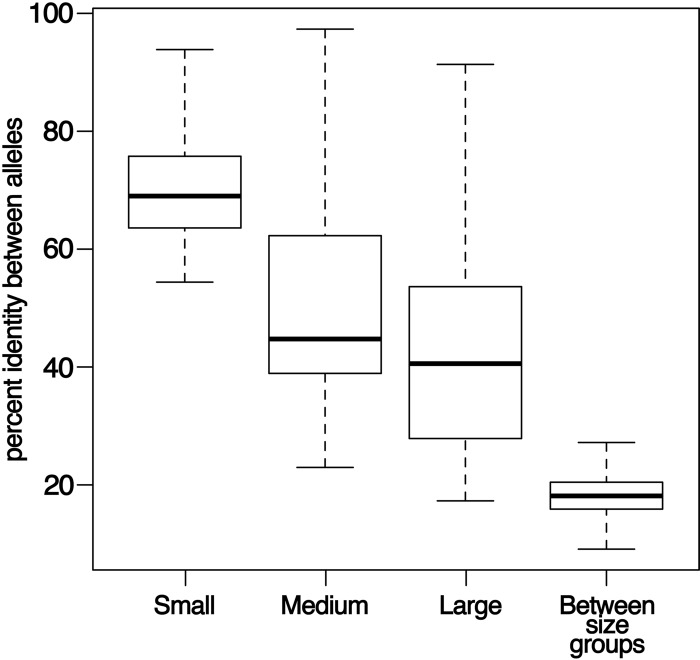
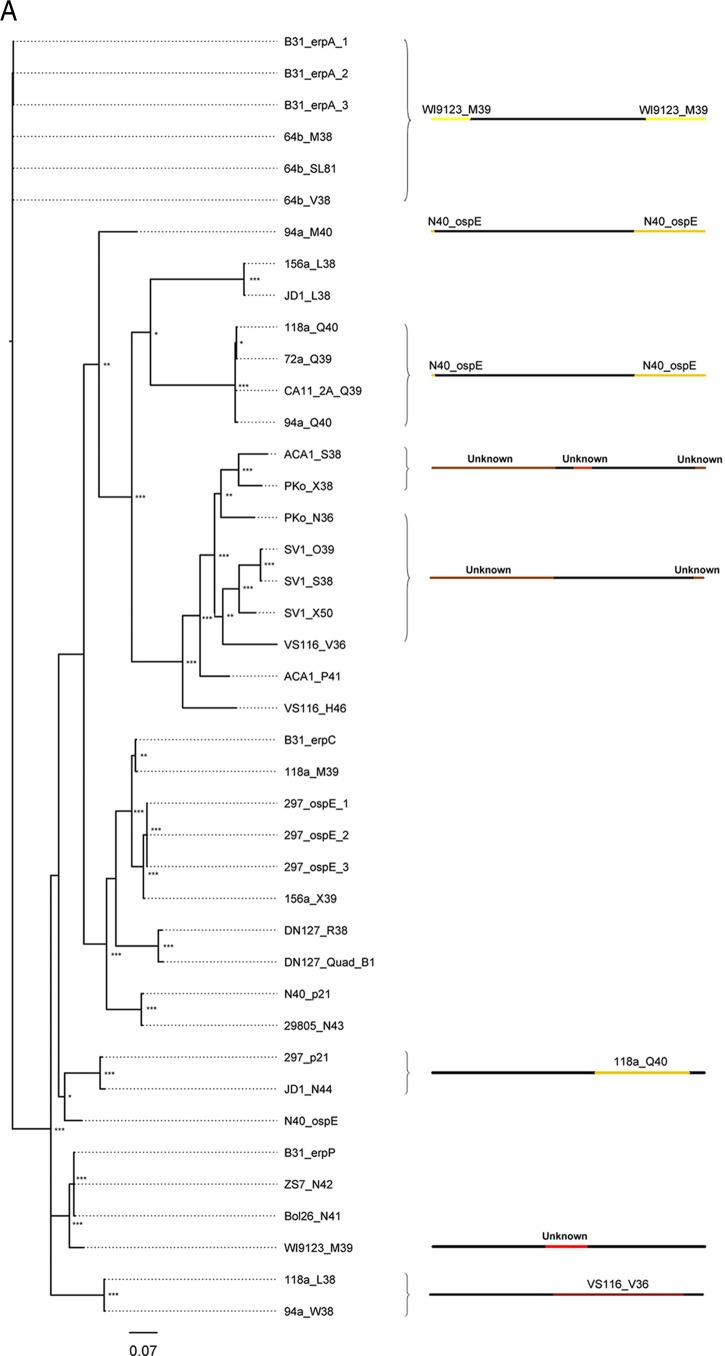
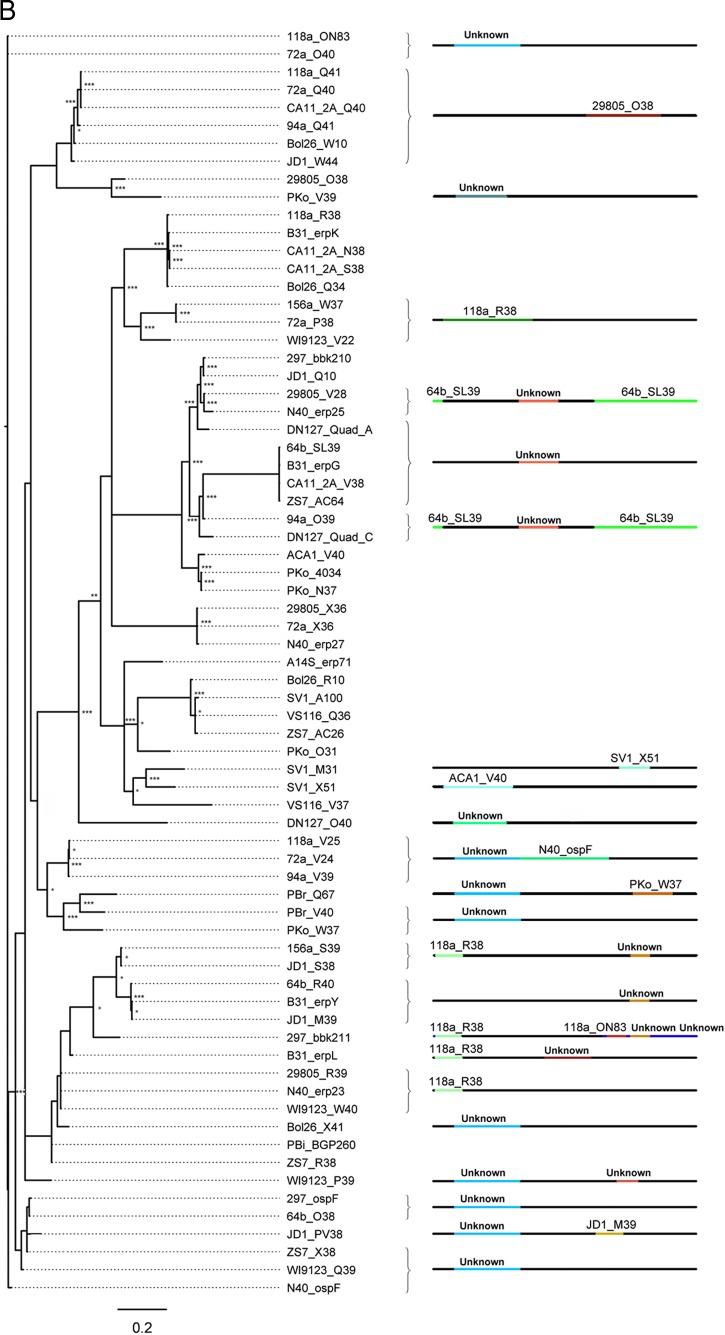
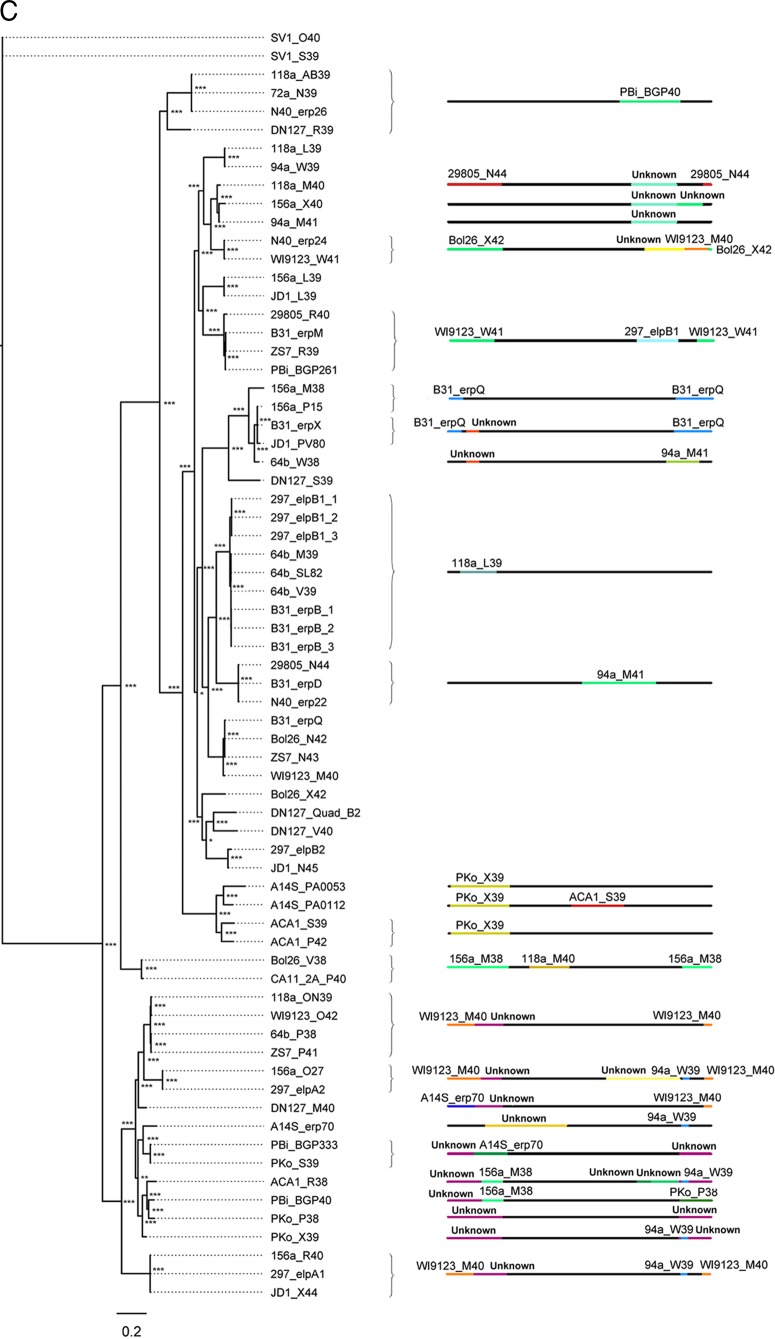
Lyme disease spirochetes possess complex genomes, consisting of a main chromosome and 20 or more smaller replicons. Among those small DNAs are the cp32 elements, a family of prophages that replicate as circular episomes. All complete cp32s contain an erp locus, which encodes surface-exposed proteins. Sequences were compared for all 193 erp alleles carried by 22 different strains of Lyme disease-causing spirochete to investigate their natural diversity and evolutionary histories. These included multiple isolates from a focus where Lyme disease is endemic in the northeastern United States and isolates from across North America and Europe. Bacteria were derived from diseased humans and from vector ticks and included members of 5 different Borrelia genospecies. All erp operon 5'-noncoding regions were found to be highly conserved, as were the initial 70 to 80 bp of all erp open reading frames, traits indicative of a common evolutionary origin. However, the majority of the protein-coding regions are highly diverse, due to numerous intra- and intergenic recombination events. Most erp alleles are chimeras derived from sequences of closely related and distantly related erp sequences and from unknown origins. Since known functions of Erp surface proteins involve interactions with various host tissue components, this diversity may reflect both their multiple functions and the abilities of Lyme disease-causing spirochetes to successfully infect a wide variety of vertebrate host species.
Figures



Similar articles
-
Erp and Rev Adhesins of the Lyme Disease Spirochete's Ubiquitous cp32 Prophages Assist the Bacterium during Vertebrate Infection.Infect Immun. 2023 Mar 15;91(3):e0025022. doi: 10.1128/iai.00250-22. Epub 2023 Feb 28. Infect Immun. 2023. PMID: 36853019 Free PMC article. Review.
-
Intra- and interbacterial genetic exchange of Lyme disease spirochete erp genes generates sequence identity amidst diversity.J Mol Evol. 2003 Sep;57(3):309-24. doi: 10.1007/s00239-003-2482-x. J Mol Evol. 2003. PMID: 14629041
-
Borrelia burgdorferi EbfC, a novel, chromosomally encoded protein, binds specific DNA sequences adjacent to erp loci on the spirochete's resident cp32 prophages.J Bacteriol. 2006 Jun;188(12):4331-9. doi: 10.1128/JB.00005-06. J Bacteriol. 2006. PMID: 16740939 Free PMC article.
-
Borrelia burgdorferi lacking all cp32 prophage plasmids retains full infectivity in mice.EMBO Rep. 2025 Apr;26(8):1997-2012. doi: 10.1038/s44319-025-00378-9. Epub 2025 Mar 19. EMBO Rep. 2025. PMID: 40108404 Free PMC article.
-
Genetics of Borrelia burgdorferi.Annu Rev Genet. 2012;46:515-36. doi: 10.1146/annurev-genet-011112-112140. Epub 2012 Sep 4. Annu Rev Genet. 2012. PMID: 22974303 Free PMC article. Review.
Cited by
-
Cas9-mediated endogenous plasmid loss in Borrelia burgdorferi.PLoS One. 2022 Nov 28;17(11):e0278151. doi: 10.1371/journal.pone.0278151. eCollection 2022. PLoS One. 2022. PMID: 36441794 Free PMC article.
-
Gene bb0318 Is Critical for the Oxidative Stress Response and Infectivity of Borrelia burgdorferi.Infect Immun. 2016 Oct 17;84(11):3141-3151. doi: 10.1128/IAI.00430-16. Print 2016 Nov. Infect Immun. 2016. PMID: 27550932 Free PMC article.
-
Erp and Rev Adhesins of the Lyme Disease Spirochete's Ubiquitous cp32 Prophages Assist the Bacterium during Vertebrate Infection.Infect Immun. 2023 Mar 15;91(3):e0025022. doi: 10.1128/iai.00250-22. Epub 2023 Feb 28. Infect Immun. 2023. PMID: 36853019 Free PMC article. Review.
-
The RpoS Gatekeeper in Borrelia burgdorferi: An Invariant Regulatory Scheme That Promotes Spirochete Persistence in Reservoir Hosts and Niche Diversity.Front Microbiol. 2019 Aug 21;10:1923. doi: 10.3389/fmicb.2019.01923. eCollection 2019. Front Microbiol. 2019. PMID: 31507550 Free PMC article.
-
Lyme Disease in Humans.Curr Issues Mol Biol. 2021;42:333-384. doi: 10.21775/cimb.042.333. Epub 2020 Dec 11. Curr Issues Mol Biol. 2021. PMID: 33303701 Free PMC article.
References
-
- Fraser CM, Casjens S, Huang WM, Sutton GG, Clayton R, Lathigra R, White O, Ketchum KA, Dodson R, Hickey EK, Gwinn M, Dougherty B, Tomb J-F, Fleischmann RD, Richardson D, Peterson J, Kerlavage AR, Quackenbush J, Salzberg S, Hanson M, van Vugt R, Palmer N, Adams MD, Gocayne J, Weidmann J, Utterback T, Watthey L, McDonald L, Artiach P, Bowman C, Garland S, Fujii C, Cotton MD, Horst K, Roberts K, Hatch B, Smith HO, Venter JC. 1997. Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi. Nature 390:580–586 - PubMed
-
- Casjens S, Palmer N, van Vugt R, Huang WM, Stevenson B, Rosa P, Lathigra R, Sutton G, Peterson J, Dodson RJ, Haft D, Hickey E, Gwinn M, White O, Fraser C. 2000. A bacterial genome in flux: the twelve linear and nine circular extrachromosomal DNAs of an infectious isolate of the Lyme disease spirochete Borrelia burgdorferi. Mol. Microbiol. 35:490–516 - PubMed
-
- Casjens SR, Eggers CH, Schwartz I. 2010. Borrelia genomics: chromosome, plasmids, bacteriophages and genetic variation, p 27–54 In Samuels DS, Radolf JD. (ed), Borrelia: molecular biology, host interaction and pathogenesis. Caister Academic Press, Norfolk, United Kingdom
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
