

Regulation of fatty acid metabolism by mTOR in adult murine hearts occurs independently of changes in PGC-1α
- PMID: 23624629
- PMCID: PMC3727103
- DOI: 10.1152/ajpheart.00877.2012
Regulation of fatty acid metabolism by mTOR in adult murine hearts occurs independently of changes in PGC-1α
Abstract
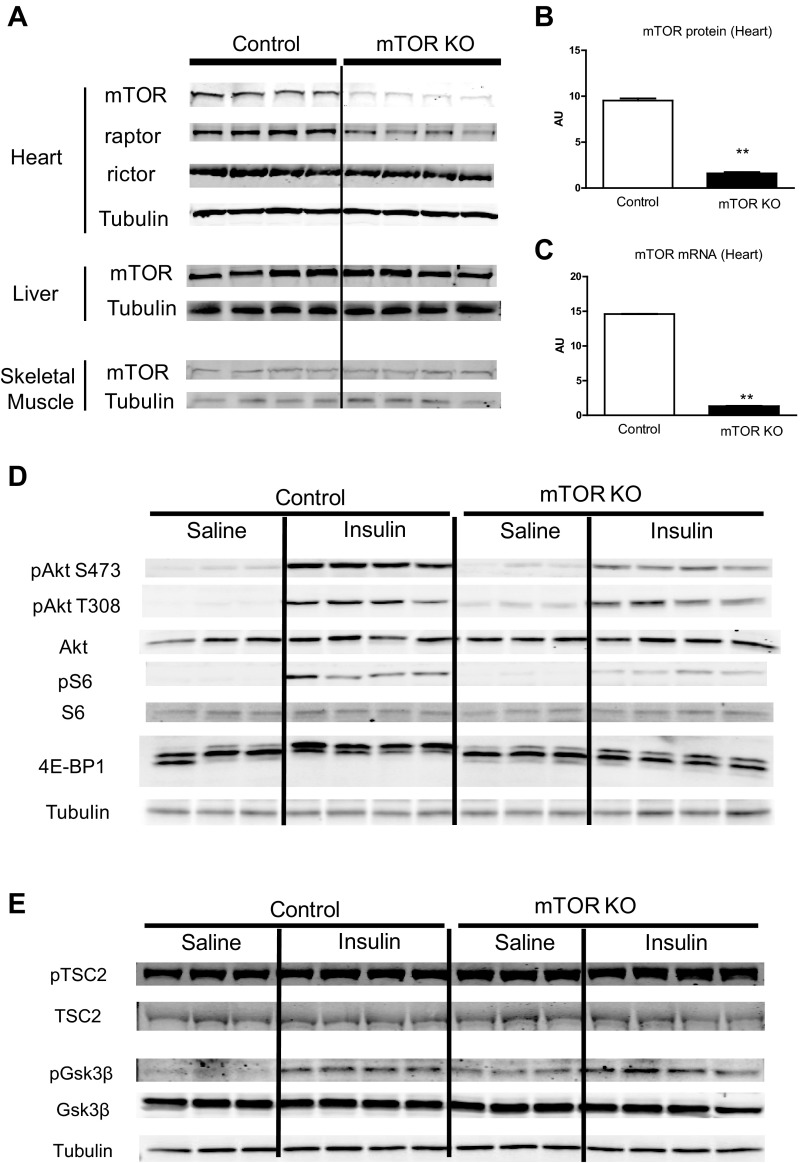
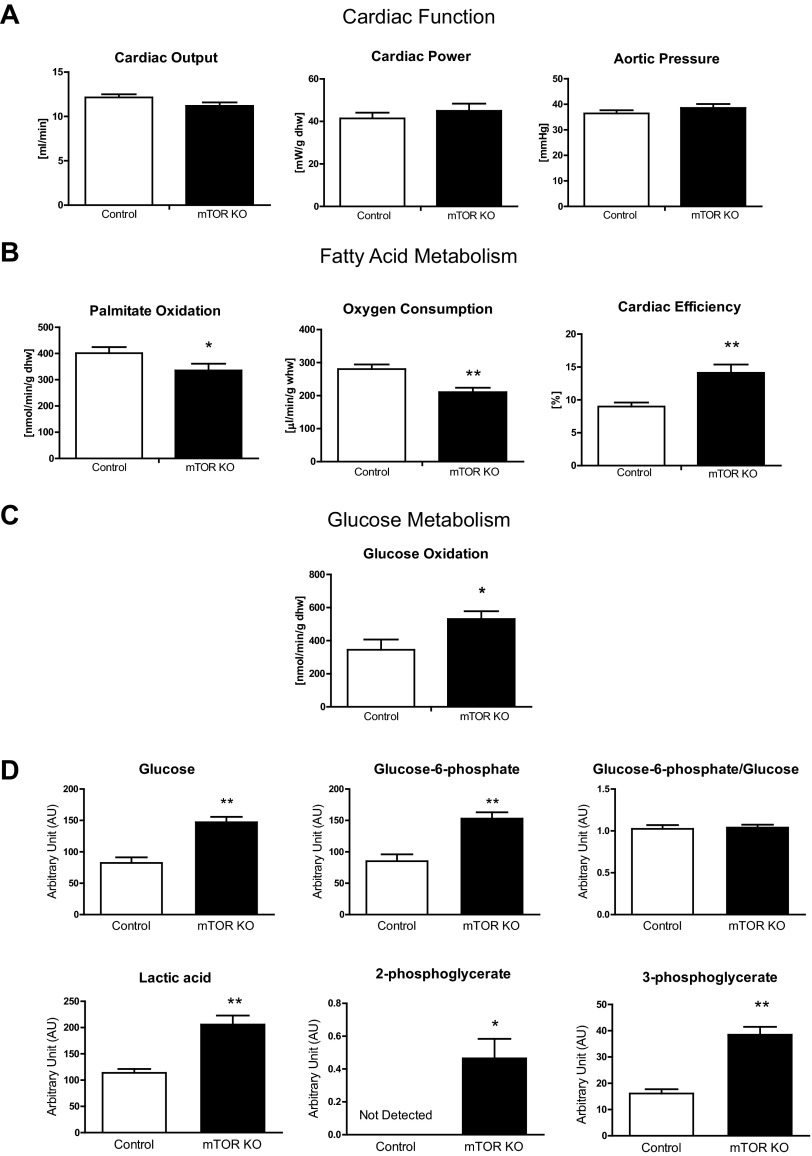
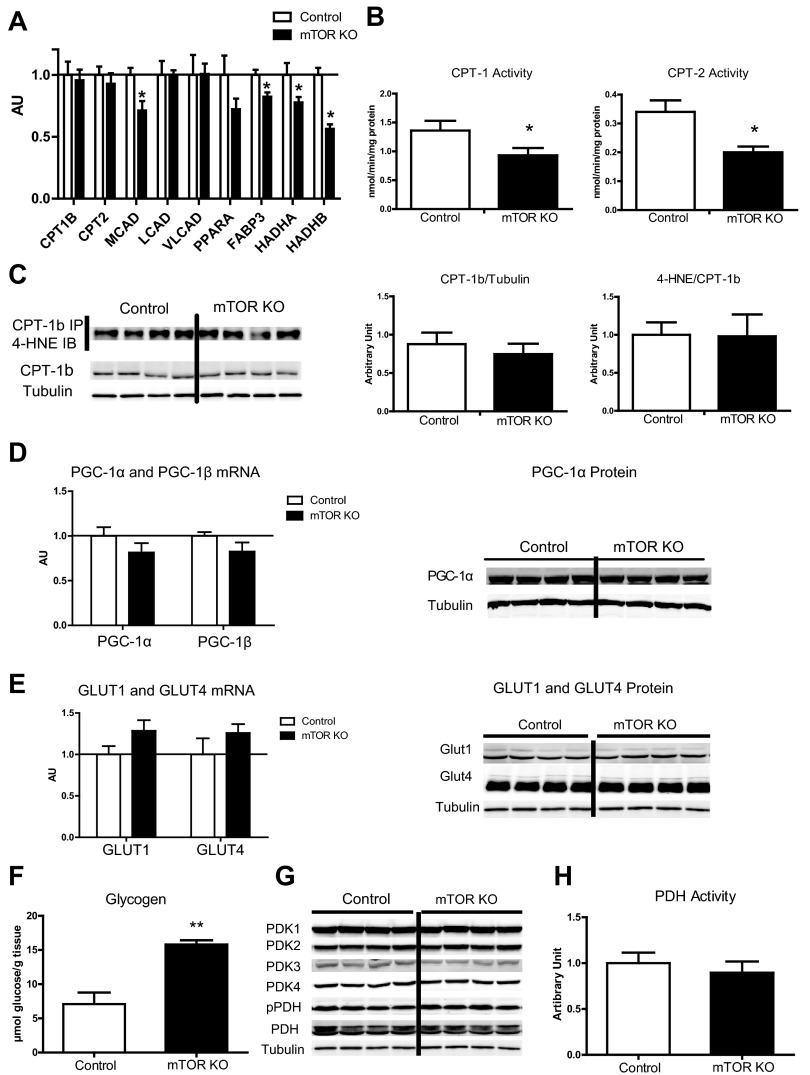
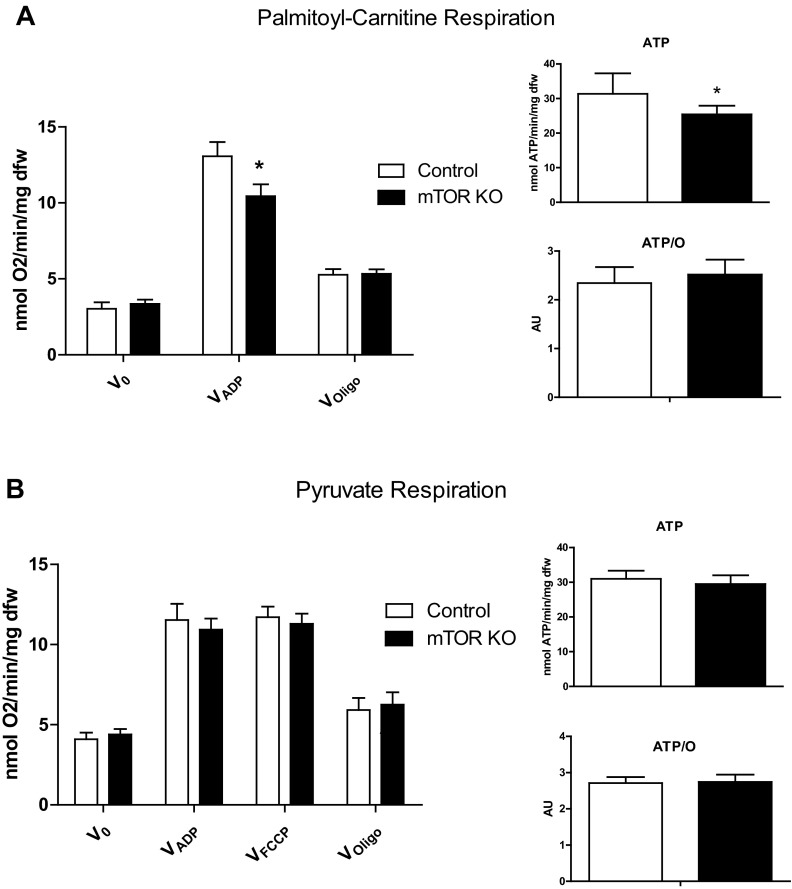
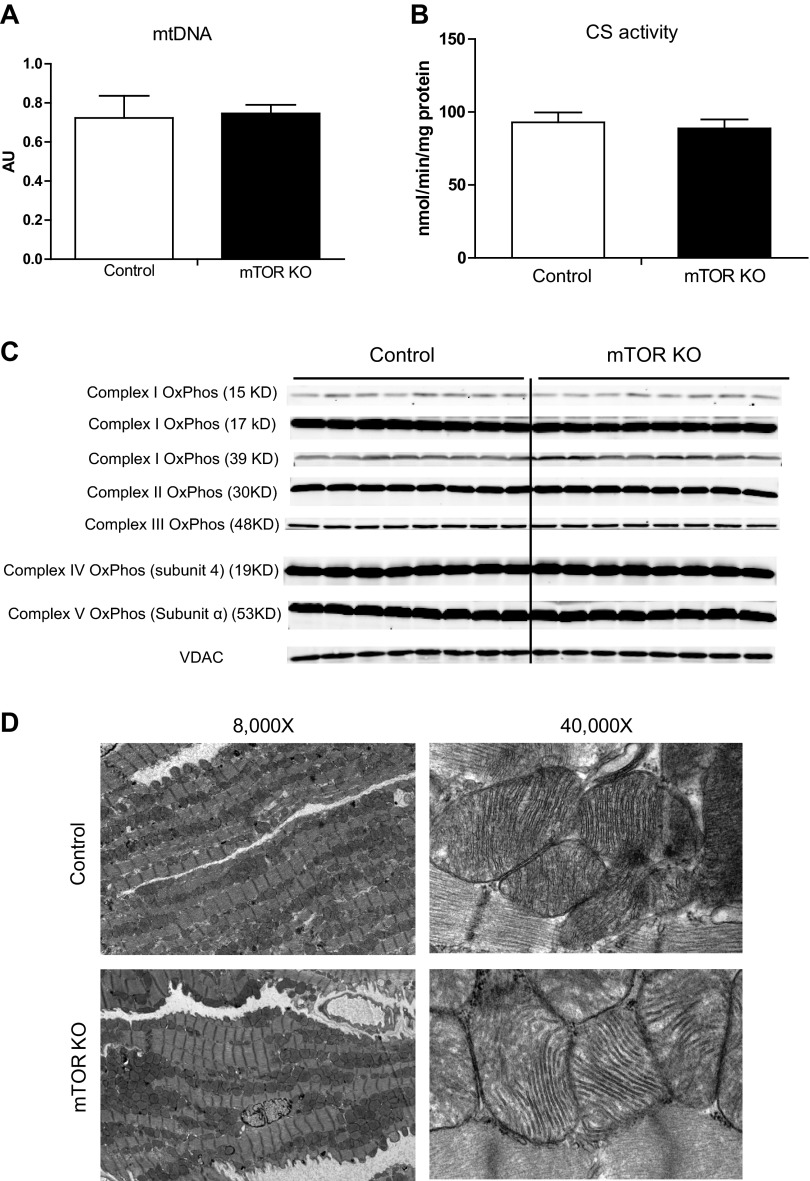
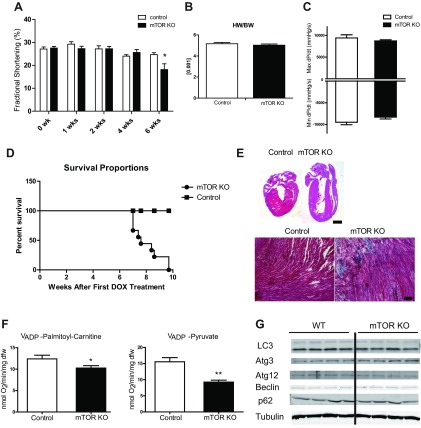
Mechanistic target of rapamycin (mTOR) is essential for cardiac development, growth, and function, but the role of mTOR in the regulation of cardiac metabolism and mitochondrial respiration is not well established. This study sought to determine cardiac metabolism and mitochondrial bioenergetics in mice with inducible deletion of mTOR in the adult heart. Doxycycline-inducible and cardiac-specific mTOR-deficient mice were generated by crossing cardiac-specific doxycycline-inducible tetO-Cre mice with mice harboring mTOR floxed alleles. Deletion of mTOR reduced mTORC1 and mTORC2 signaling after in vivo insulin stimulation. Maximum and minimum dP/dt measured by cardiac catheterization in vivo under anesthesia and cardiac output, cardiac power, and aortic pressure in ex vivo working hearts were unchanged, suggesting preserved cardiac function 4 wk after doxycycline treatment. However, myocardial palmitate oxidation was impaired, whereas glucose oxidation was increased. Consistent with reduced palmitate oxidation, expression of fatty acid metabolism genes fatty acid-binding protein 3, medium-chain acyl-CoA dehydrogenase, and hydroxyacyl-CoA dehydrogenase/3-ketoacyl-CoA thiolase/enoyl-CoA hydratase (trifunctional protein)-α and -β was reduced, and carnitine palmitoyl transferase-1 and -2 enzymatic activity was decreased. Mitochondrial palmitoyl carnitine respiration was diminished. However, mRNA for peroxisome proliferator-activated receptor-γ coactivator (PGC)-1α and -1β, protein levels of PGC-1α, and electron transport chain subunits, mitochondrial DNA, and morphology were unchanged. Also, pyruvate-supported and FCCP-stimulated respirations were unchanged, suggesting that mTOR deletion induces a specific defect in fatty acid utilization. In conclusion, mTOR regulates mitochondrial fatty acid utilization but not glucose utilization in the heart via mechanisms that are independent of changes in PGC expression.
Keywords: cardiac substrate metabolism; mechanistic target of rapamycin; mitochondrial respiration; peroxisome proliferator-activated receptor-γ coactivator-1α.
Figures
Similar articles
-
The transcriptional coactivator PGC-1alpha is essential for maximal and efficient cardiac mitochondrial fatty acid oxidation and lipid homeostasis.Am J Physiol Heart Circ Physiol. 2008 Jul;295(1):H185-96. doi: 10.1152/ajpheart.00081.2008. Epub 2008 May 16. Am J Physiol Heart Circ Physiol. 2008. PMID: 18487436 Free PMC article.
-
Myocardial infarction in rats causes partial impairment in insulin response associated with reduced fatty acid oxidation and mitochondrial gene expression.J Thorac Cardiovasc Surg. 2010 Nov;140(5):1160-7. doi: 10.1016/j.jtcvs.2010.08.003. Epub 2010 Sep 17. J Thorac Cardiovasc Surg. 2010. PMID: 20850803
-
Induction of heart failure by minimally invasive aortic constriction in mice: reduced peroxisome proliferator-activated receptor γ coactivator levels and mitochondrial dysfunction.J Thorac Cardiovasc Surg. 2011 Feb;141(2):492-500, 500.e1. doi: 10.1016/j.jtcvs.2010.03.029. Epub 2010 May 5. J Thorac Cardiovasc Surg. 2011. PMID: 20447656
-
Regulation of energy metabolism by long-chain fatty acids.Prog Lipid Res. 2014 Jan;53:124-44. doi: 10.1016/j.plipres.2013.12.001. Epub 2013 Dec 18. Prog Lipid Res. 2014. PMID: 24362249 Review.
-
Nuclear receptor signaling and cardiac energetics.Circ Res. 2004 Sep 17;95(6):568-78. doi: 10.1161/01.RES.0000141774.29937.e3. Circ Res. 2004. PMID: 15375023 Review.
Cited by
-
mTOR coordinates protein synthesis, mitochondrial activity and proliferation.Cell Cycle. 2015;14(4):473-80. doi: 10.4161/15384101.2014.991572. Cell Cycle. 2015. PMID: 25590164 Free PMC article. Review.
-
Mitochondrial and Metabolic Gene Expression in the Aged Rat Heart.Front Physiol. 2016 Aug 23;7:352. doi: 10.3389/fphys.2016.00352. eCollection 2016. Front Physiol. 2016. PMID: 27601998 Free PMC article.
-
Loss of long-chain acyl-CoA synthetase isoform 1 impairs cardiac autophagy and mitochondrial structure through mechanistic target of rapamycin complex 1 activation.FASEB J. 2015 Nov;29(11):4641-53. doi: 10.1096/fj.15-272732. Epub 2015 Jul 28. FASEB J. 2015. PMID: 26220174 Free PMC article.
-
Mammalian target of rapamycin signaling in cardiac physiology and disease.Circ Res. 2014 Jan 31;114(3):549-64. doi: 10.1161/CIRCRESAHA.114.302022. Circ Res. 2014. PMID: 24481845 Free PMC article. Review.
-
Mammalian Target of Rapamycin: A Metabolic Rheostat for Regulating Adipose Tissue Function and Cardiovascular Health.Am J Pathol. 2019 Mar;189(3):492-501. doi: 10.1016/j.ajpath.2018.11.013. Am J Pathol. 2019. PMID: 30803496 Free PMC article. Review.
References
-
- Bentzinger CF, Romanino K, Cloëtta D, Lin S, Mascarenhas JB, Oliveri F, Xia J, Casanova E, Costa CF, Brink M, Zorzato F, Hall MN, Rüegg MA. Skeletal muscle-specific ablation of raptor, but not of rictor, causes metabolic changes and results in muscle dystrophy. Cell Metab 8: 411– 424, 2008 - PubMed
-
- Boudina S, Sena S, O'Neill BT, Tathireddy P, Young ME, Abel ED. Reduced mitochondrial oxidative capacity and increased mitochondrial uncoupling impair myocardial energetics in obesity. Circulation 112: 2686– 2695, 2005 - PubMed
-
- Boudina S, Sena S, Theobald H, Sheng X, Wright JJ, Hu XX, Aziz S, Johnson JI, Bugger H, Zaha VG, Abel ED. Mitochondrial energetics in the heart in obesity-related diabetes: direct evidence for increased uncoupled respiration and activation of uncoupling proteins. Diabetes 56: 2457– 2466, 2007 - PubMed
-
- Buchanan J, Mazumder PK, Hu P, Chakrabarti G, Roberts MW, Yun UJ, Cooksey RC, Litwin SE, Abel ED. Reduced cardiac efficiency and altered substrate metabolism precedes the onset of hyperglycemia and contractile dysfunction in two mouse models of insulin resistance and obesity. Endocrinology 146: 5341– 5349, 2005 - PubMed
-
- Cunningham JT, Rodgers JT, Arlow DH, Vazquez F, Mootha VK, Puigserver P. mTOR controls mitochondrial oxidative function through a YY1-PGC-1α transcriptional complex. Nature 450: 736– 740, 2007 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous