

Reduced tonoplast fast-activating and slow-activating channel activity is essential for conferring salinity tolerance in a facultative halophyte, quinoa
- PMID: 23624857
- PMCID: PMC3668082
- DOI: 10.1104/pp.113.216572
Reduced tonoplast fast-activating and slow-activating channel activity is essential for conferring salinity tolerance in a facultative halophyte, quinoa
Abstract
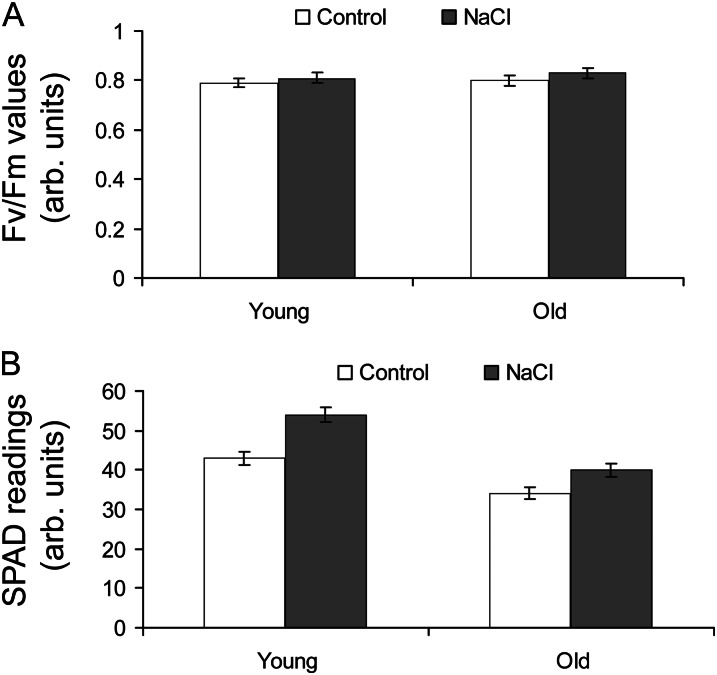
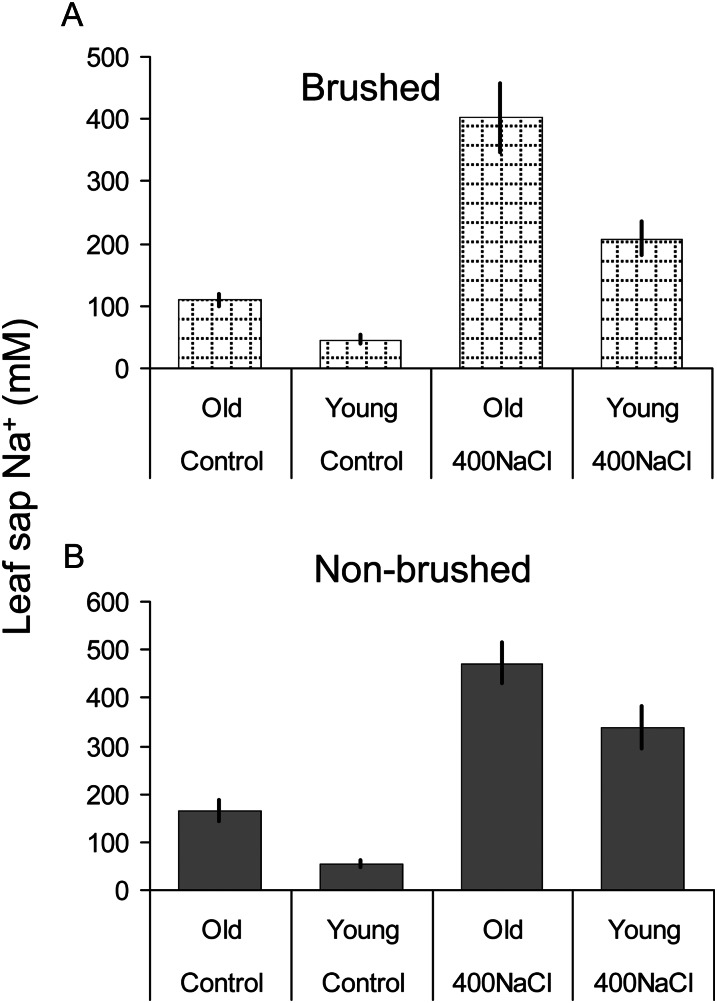
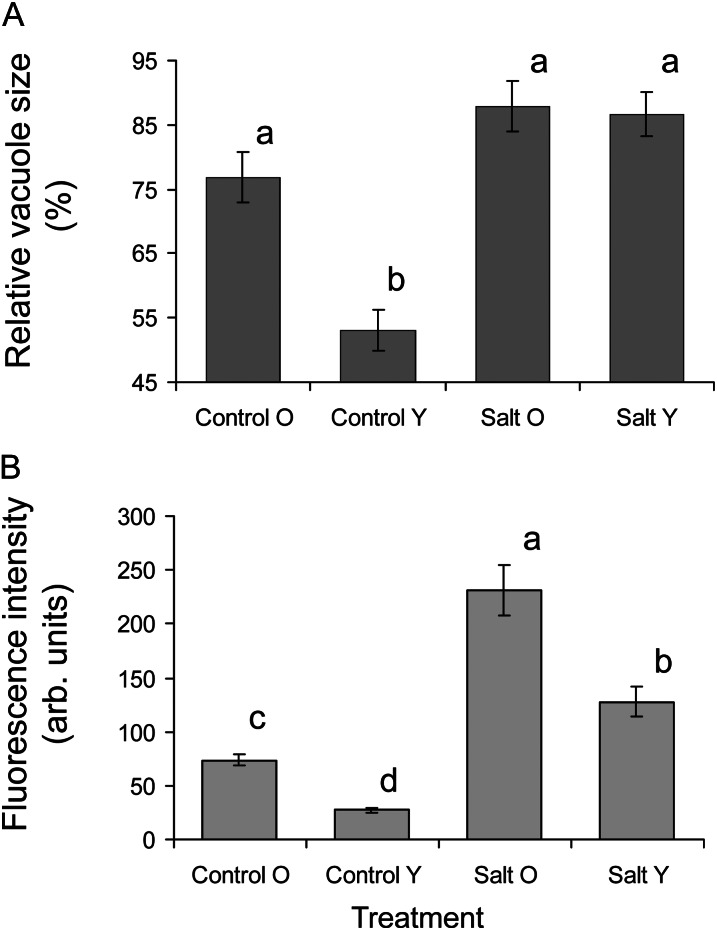
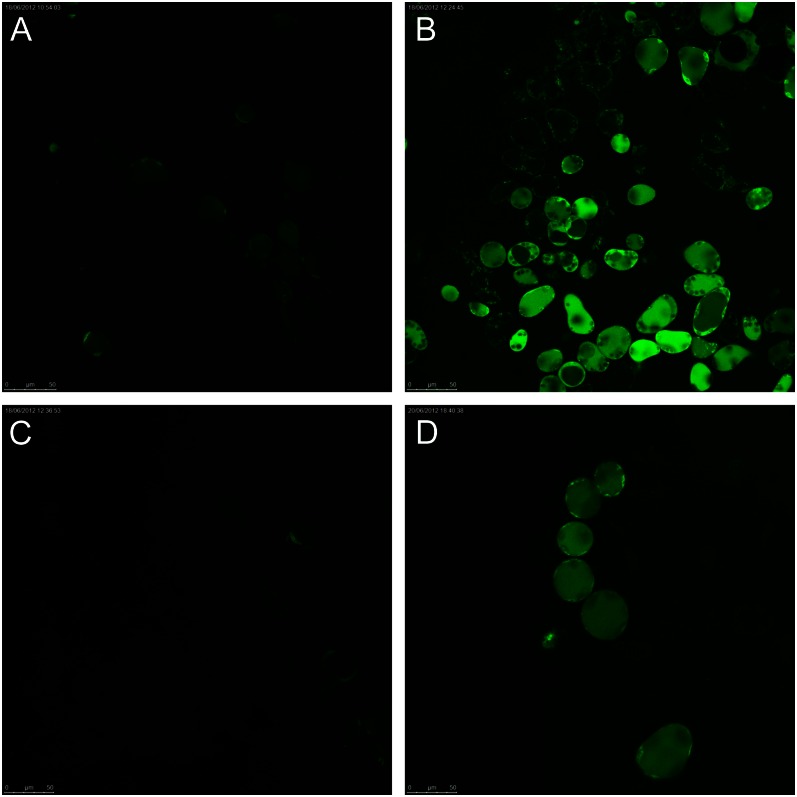
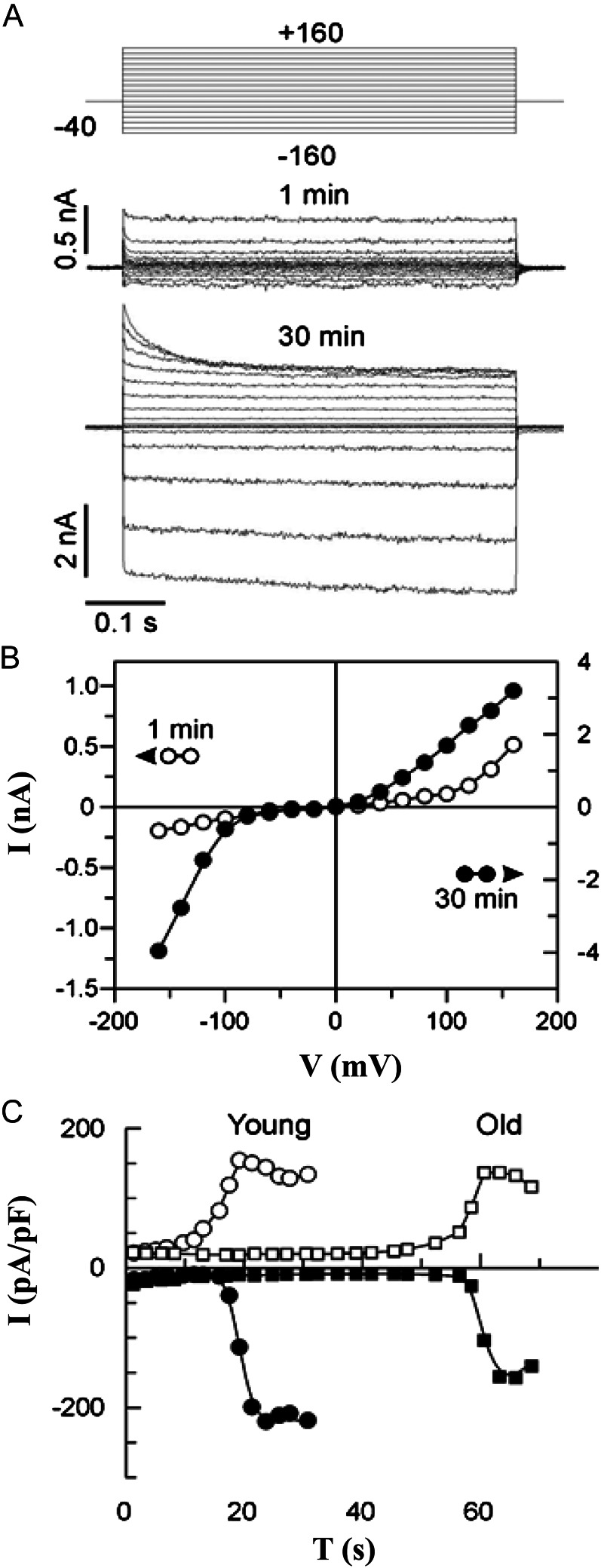
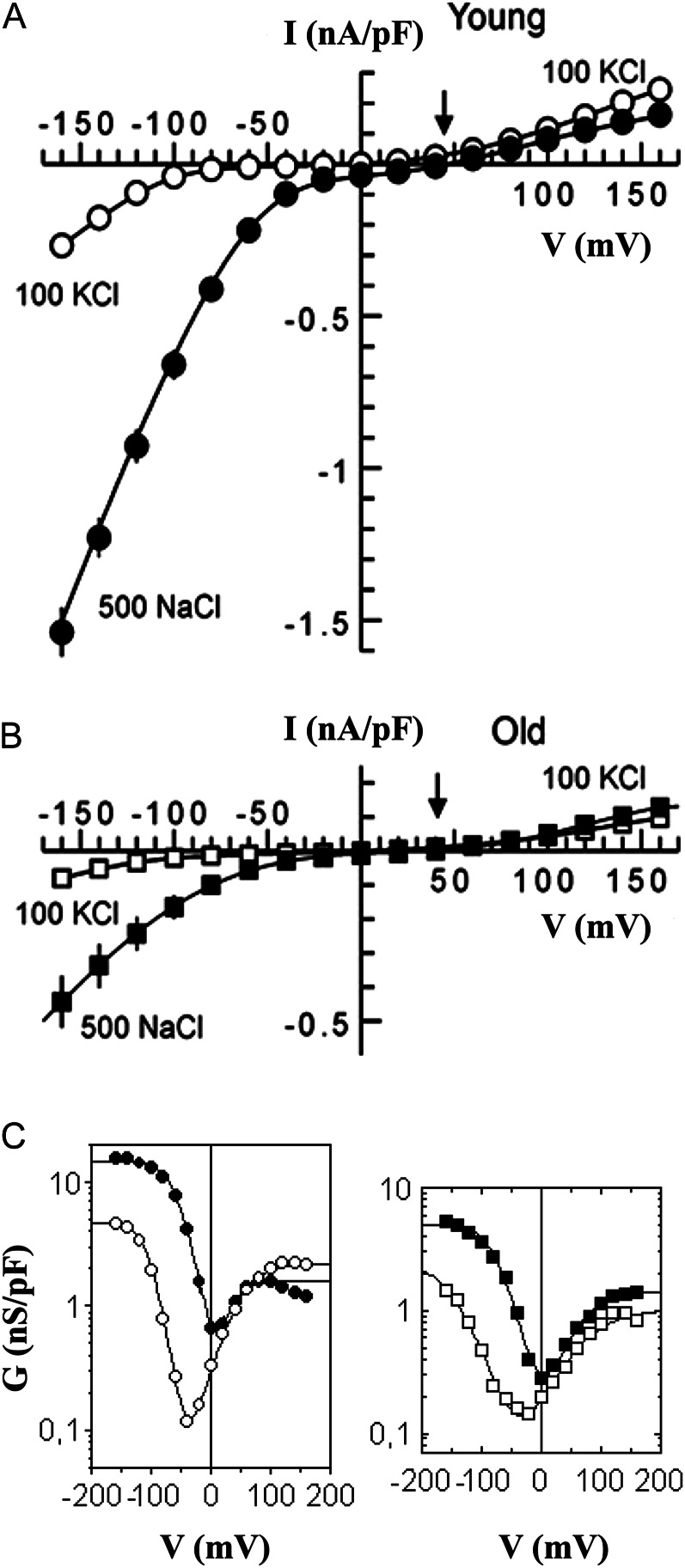
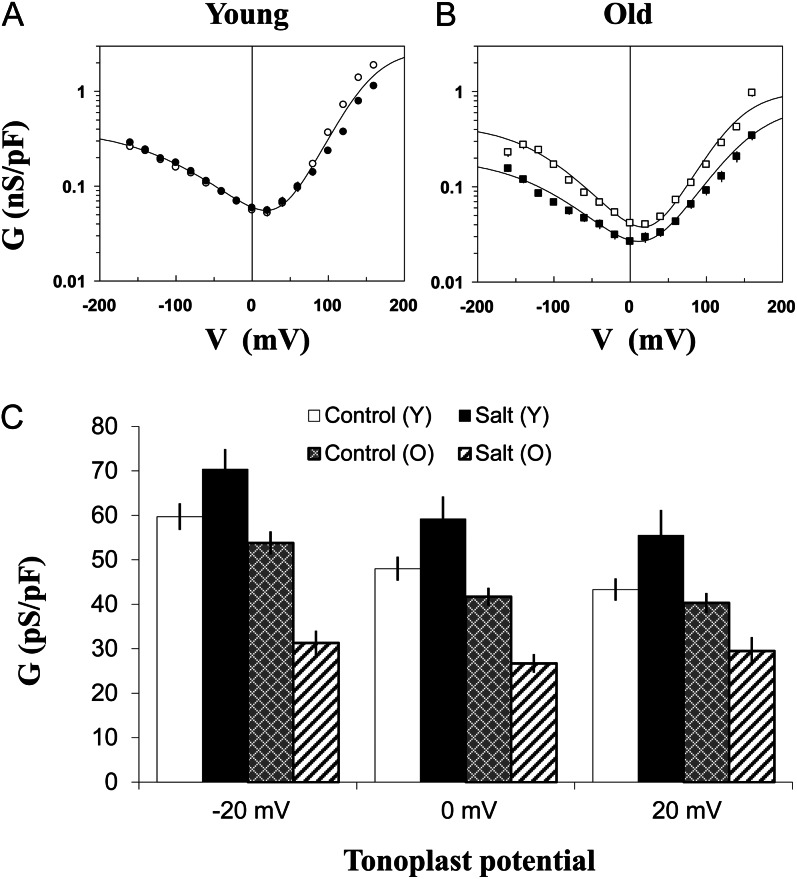
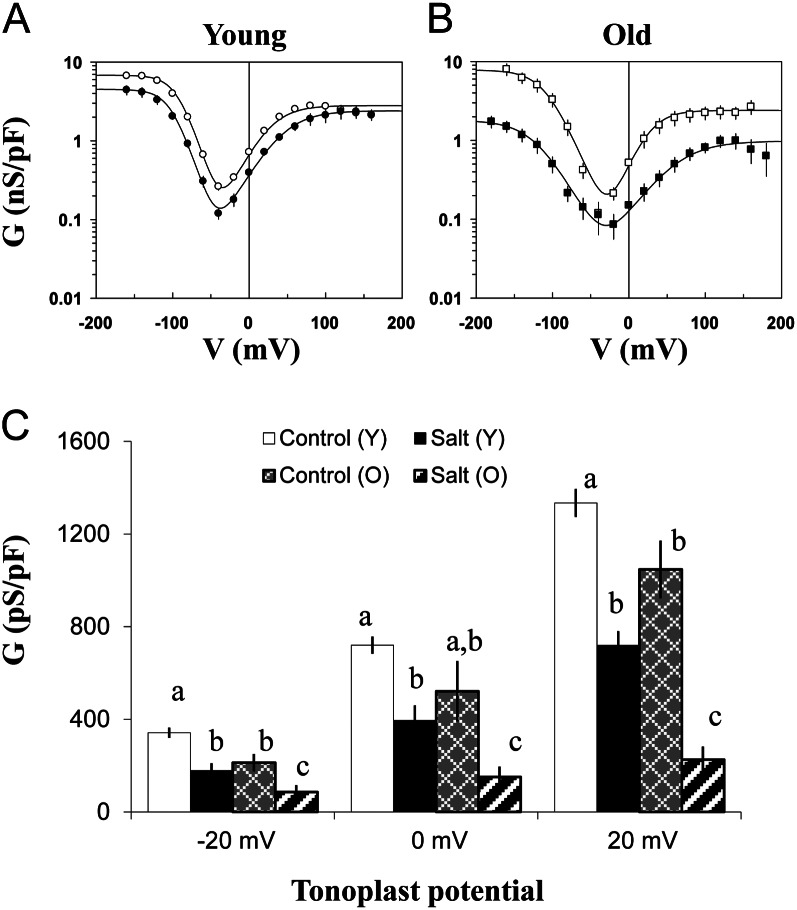
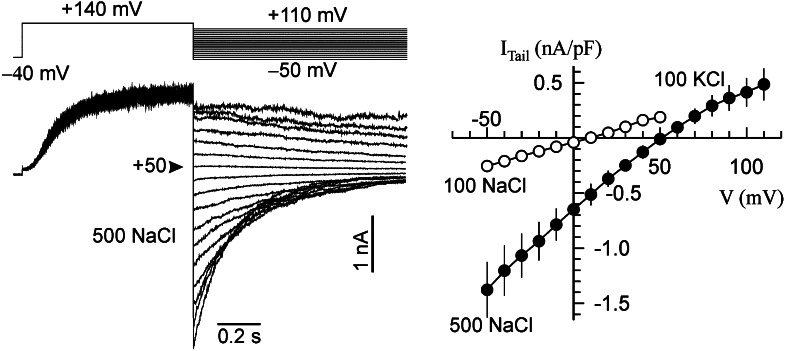
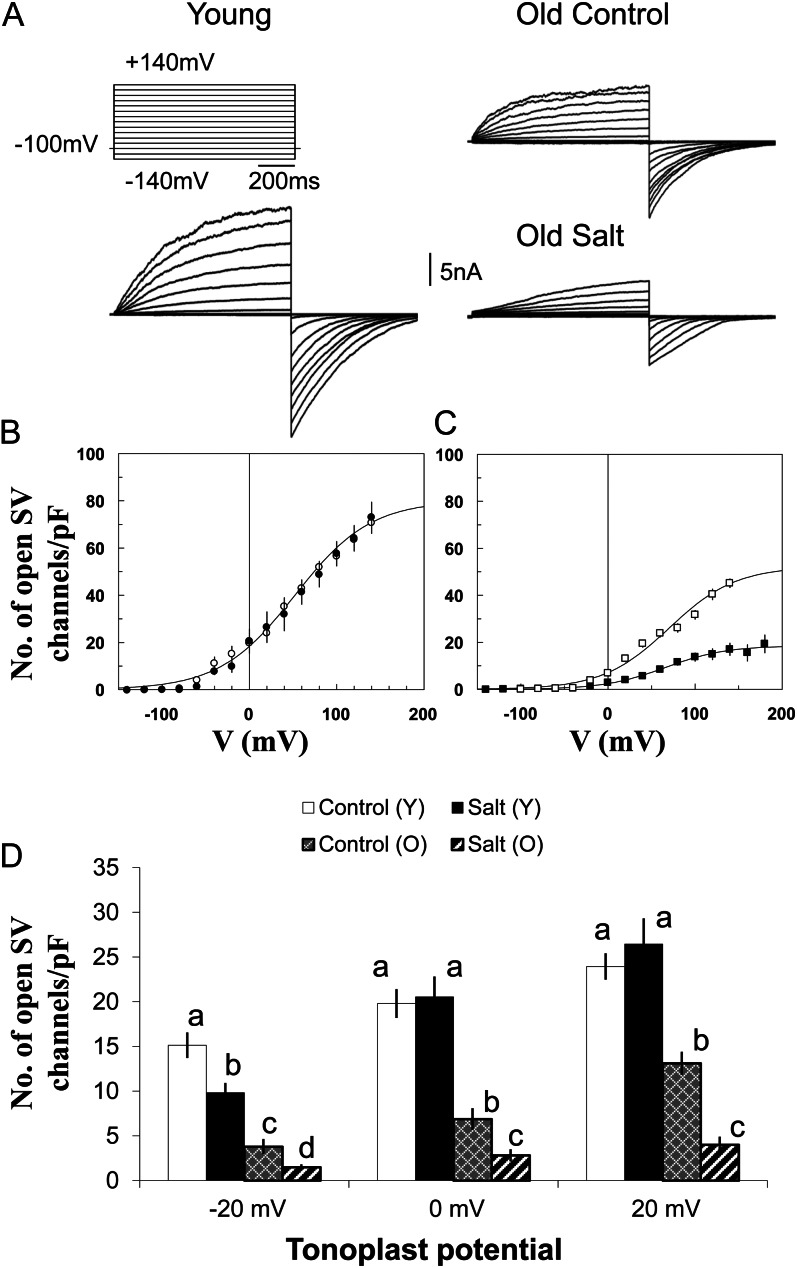
Halophyte species implement a "salt-including" strategy, sequestering significant amounts of Na(+) to cell vacuoles. This requires a reduction of passive Na(+) leak from the vacuole. In this work, we used quinoa (Chenopodium quinoa) to investigate the ability of halophytes to regulate Na(+)-permeable slow-activating (SV) and fast-activating (FV) tonoplast channels, linking it with Na(+) accumulation in mesophyll cells and salt bladders as well as leaf photosynthetic efficiency under salt stress. Our data indicate that young leaves rely on Na(+) exclusion to salt bladders, whereas old ones, possessing far fewer salt bladders, depend almost exclusively on Na(+) sequestration to mesophyll vacuoles. Moreover, although old leaves accumulate more Na(+), this does not compromise their leaf photochemistry. FV and SV channels are slightly more permeable for K(+) than for Na(+), and vacuoles in young leaves express less FV current and with a density unchanged in plants subjected to high (400 mm NaCl) salinity. In old leaves, with an intrinsically lower density of the FV current, FV channel density decreases about 2-fold in plants grown under high salinity. In contrast, intrinsic activity of SV channels in vacuoles from young leaves is unchanged under salt stress. In vacuoles of old leaves, however, it is 2- and 7-fold lower in older compared with young leaves in control- and salt-grown plants, respectively. We conclude that the negative control of SV and FV tonoplast channel activity in old leaves reduces Na(+) leak, thus enabling efficient sequestration of Na(+) to their vacuoles. This enables optimal photosynthetic performance, conferring salinity tolerance in quinoa species.
Figures

References
-
- Adams P, Nelson D, Yamada S, Chmara W, Jensen RG, Bohnert HJ, Griffiths H. (1998) Growth and development of Mesembryanthemum crystallinum (Aizoaceae). New Phytol 138: 171–190 - PubMed
-
- Adams P, Thomas JC, Vernon DM, Bohnert HJ, Jensen RG. (1992) Distinct cellular and organismic responses to salt stress. Plant Cell Physiol 33: 1215–1223
-
- Adolf VI, Shabala S, Andersen MN, Razzaghi F, Jacobsen S-E. (2012) Varietal differences of quinoa’s tolerance to saline conditions. Plant Soil 357: 117–129
-
- Agarie S, Shimoda T, Shimizu Y, Baumann K, Sunagawa H, Kondo A, Ueno O, Nakahara T, Nose A, Cushman JC. (2007) Salt tolerance, salt accumulation, and ionic homeostasis in an epidermal bladder-cell-less mutant of the common ice plant Mesembryanthemum crystallinum. J Exp Bot 58: 1957–1967 - PubMed
-
- Allen GJ, Amtmann A, Sanders D. (1998) Calcium-dependent and calcium independent K+ mobilization channels in Vicia faba guard cell vacuoles. J Exp Bot 49: 305–318
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
