

A novel approach to quantify G-protein-coupled receptor dimerization equilibrium using bioluminescence resonance energy transfer
- PMID: 23625495
- PMCID: PMC4091634
- DOI: 10.1007/978-1-62703-426-5_7
A novel approach to quantify G-protein-coupled receptor dimerization equilibrium using bioluminescence resonance energy transfer
Abstract
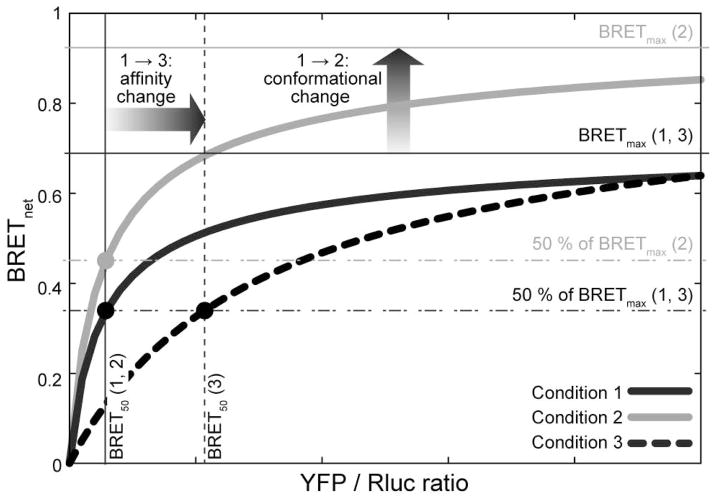
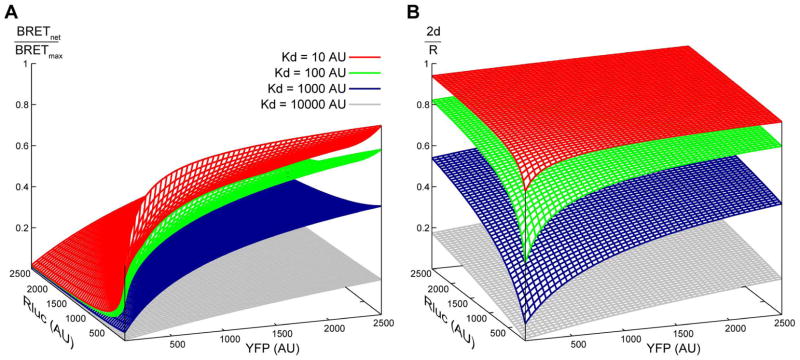
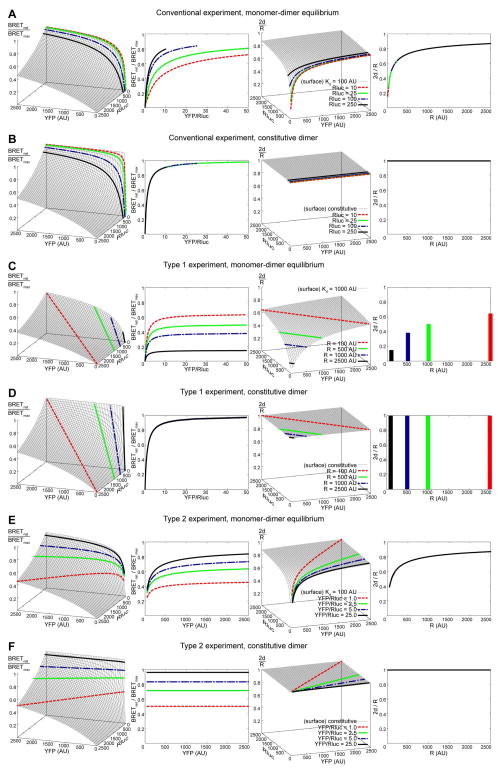
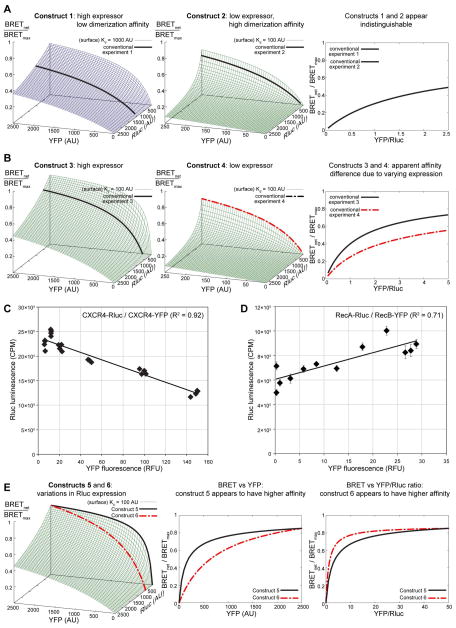
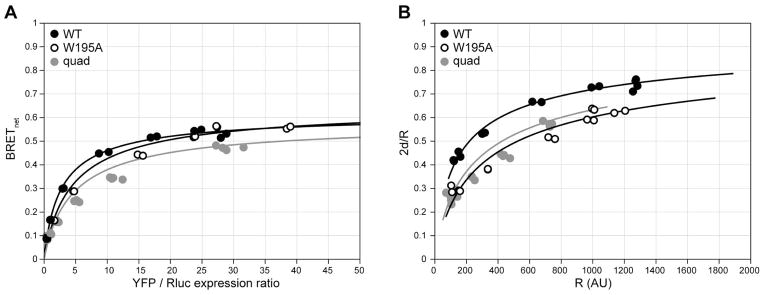
Along with other resonance energy transfer techniques, bioluminescence resonance energy transfer (BRET) has emerged as an important method for demonstrating protein-protein interactions in cells. In the field of G-protein-coupled receptors, including chemokine receptors, BRET has been widely used to investigate homo- and heterodimerization, a feature of their interactions that is emerging as integral to function and regulation. While demonstrating the existence of dimers for a given receptor proved to be fairly straightforward, quantitative comparisons of different receptors or mutants are nontrivial because of inevitable variations in the expression of receptor constructs. The uncontrollable parameters of the cellular expression machinery make amounts of transfected DNA extremely poor predictors for the expression levels of BRET donor and acceptor receptor constructs, even in relative terms. In this chapter, we show that properly accounting for receptor expression levels is critical for quantitative interpretation of BRET data. We also provide a comprehensive account of expected responses in all types of BRET experiments and propose a framework for uniform and accurate quantitative treatment of these responses. The framework allows analysis of both homodimer and heterodimer BRET data. The important caveats and obstacles for quantitative treatment are outlined, and the utility of the approach is illustrated by its application to the homodimerization of wild-type (WT) and mutant forms of the chemokine receptor CXCR4.
Figures
Similar articles
-
Improved methodical approach for quantitative BRET analysis of G Protein Coupled Receptor dimerization.PLoS One. 2014 Oct 17;9(10):e109503. doi: 10.1371/journal.pone.0109503. eCollection 2014. PLoS One. 2014. PMID: 25329164 Free PMC article.
-
Using quantitative BRET to assess G protein-coupled receptor homo- and heterodimerization.Methods Mol Biol. 2011;756:183-200. doi: 10.1007/978-1-61779-160-4_9. Methods Mol Biol. 2011. PMID: 21870226
-
Characterization of heteromeric complexes between chemokine (C-X-C motif) receptor 4 and α1-adrenergic receptors utilizing intermolecular bioluminescence resonance energy transfer assays.Biochem Biophys Res Commun. 2020 Jul 23;528(2):368-375. doi: 10.1016/j.bbrc.2020.02.094. Epub 2020 Feb 19. Biochem Biophys Res Commun. 2020. PMID: 32085899 Free PMC article.
-
Bioluminescence Resonance Energy Transfer as a Method to Study Protein-Protein Interactions: Application to G Protein Coupled Receptor Biology.Molecules. 2019 Feb 1;24(3):537. doi: 10.3390/molecules24030537. Molecules. 2019. PMID: 30717191 Free PMC article. Review.
-
Application of BRET for studying G protein-coupled receptors.Mini Rev Med Chem. 2014 May;14(5):411-25. doi: 10.2174/1389557514666140428113708. Mini Rev Med Chem. 2014. PMID: 24766382 Review.
Cited by
-
BRET evidence that β2 adrenergic receptors do not oligomerize in cells.Sci Rep. 2015 May 8;5:10166. doi: 10.1038/srep10166. Sci Rep. 2015. PMID: 25955971 Free PMC article.
-
Improved methodical approach for quantitative BRET analysis of G Protein Coupled Receptor dimerization.PLoS One. 2014 Oct 17;9(10):e109503. doi: 10.1371/journal.pone.0109503. eCollection 2014. PLoS One. 2014. PMID: 25329164 Free PMC article.
-
Chemokine receptor CXCR4 oligomerization is disrupted selectively by the antagonist ligand IT1t.J Biol Chem. 2021 Jan-Jun;296:100139. doi: 10.1074/jbc.RA120.016612. Epub 2020 Dec 6. J Biol Chem. 2021. PMID: 33268380 Free PMC article.
-
Common activation mechanism of class A GPCRs.Elife. 2019 Dec 19;8:e50279. doi: 10.7554/eLife.50279. Elife. 2019. PMID: 31855179 Free PMC article.
-
Computationally designed GPCR quaternary structures bias signaling pathway activation.Nat Commun. 2022 Nov 11;13(1):6826. doi: 10.1038/s41467-022-34382-7. Nat Commun. 2022. PMID: 36369272 Free PMC article.
References
-
- Bayburt TH, Leitz AJ, Xie G, Oprian DD, Sligar SG. Transducin activation by nanoscale lipid bilayers containing one and two rhodopsins. J Biol Chem. 2007;282:14875–14881. - PubMed
-
- Angers S, Salahpour A, Bouvier M. Dimerization: an emerging concept for G protein-coupled receptor ontogeny and function. Annu Rev Pharmacol Toxicol. 2002;42:409–435. - PubMed
-
- Springael JY, Urizar E, Parmentier M. Dimerization of chemokine receptors and its functional consequences. Cytokine Growth Factor Rev. 2005;16:611–623. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
