

ATM influences the efficiency of TCRβ rearrangement, subsequent TCRβ-dependent T cell development, and generation of the pre-selection TCRβ CDR3 repertoire
- PMID: 23626787
- PMCID: PMC3633875
- DOI: 10.1371/journal.pone.0062188
ATM influences the efficiency of TCRβ rearrangement, subsequent TCRβ-dependent T cell development, and generation of the pre-selection TCRβ CDR3 repertoire
Abstract
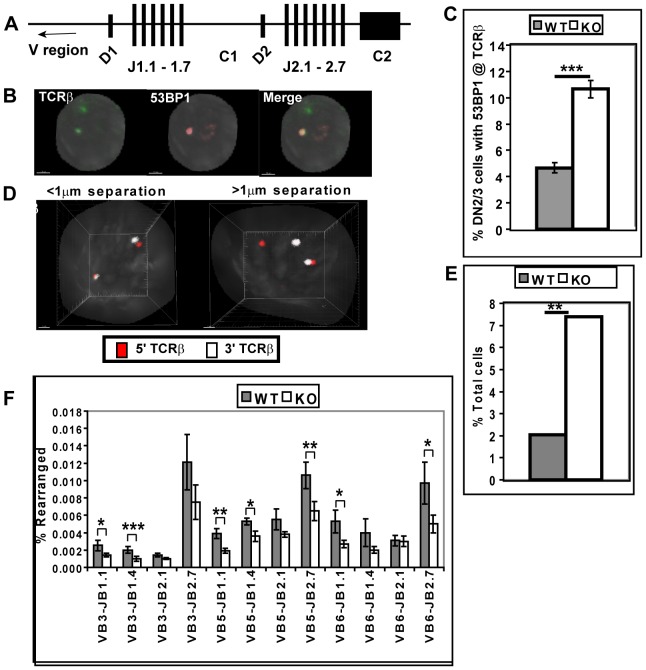
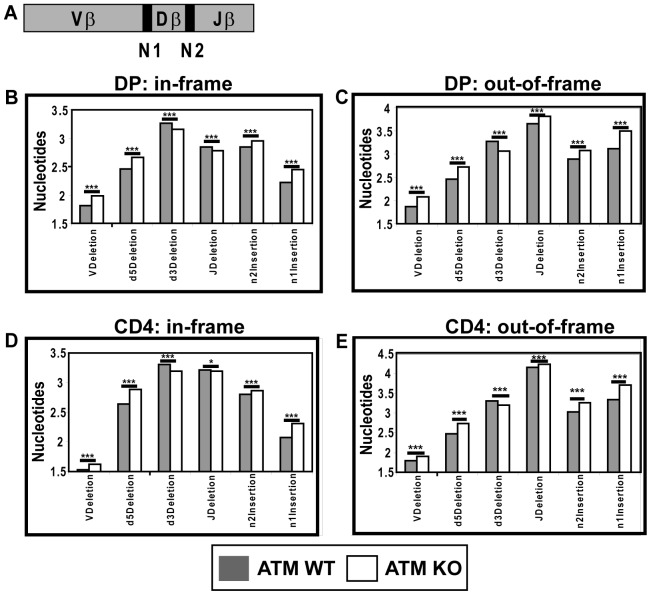
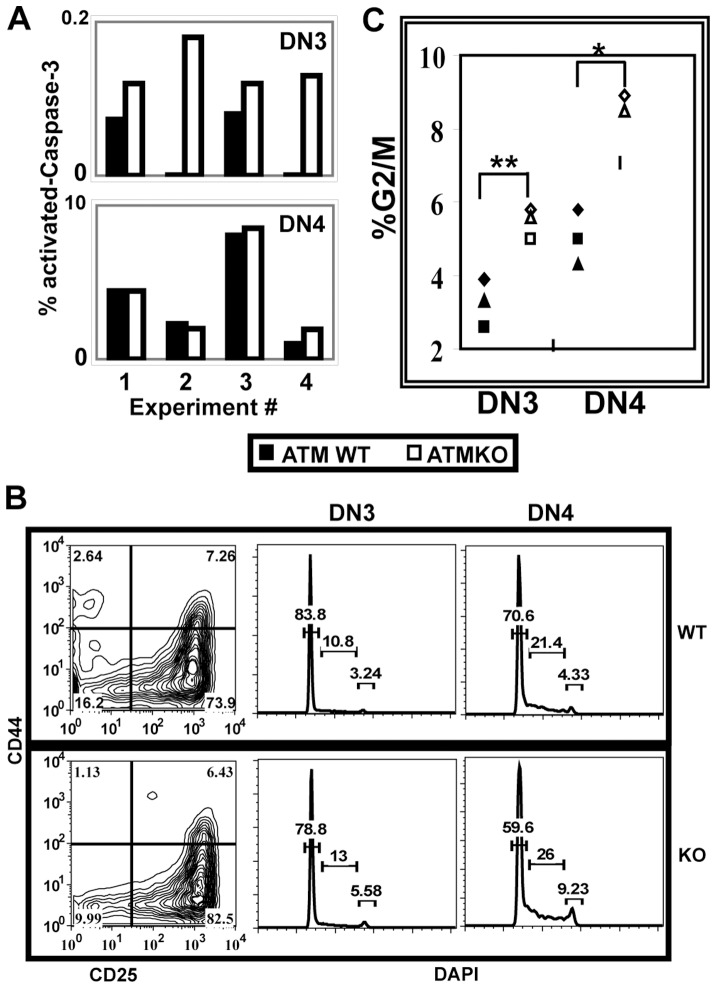
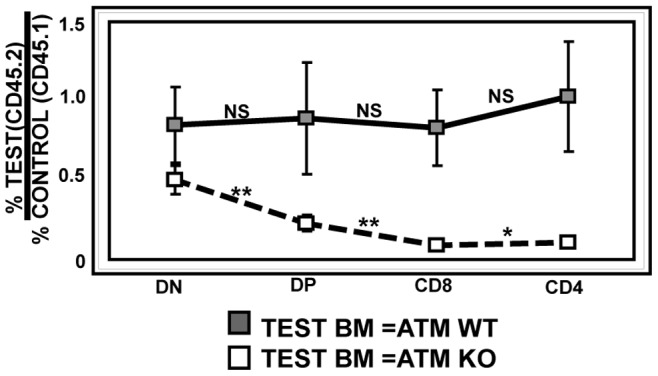
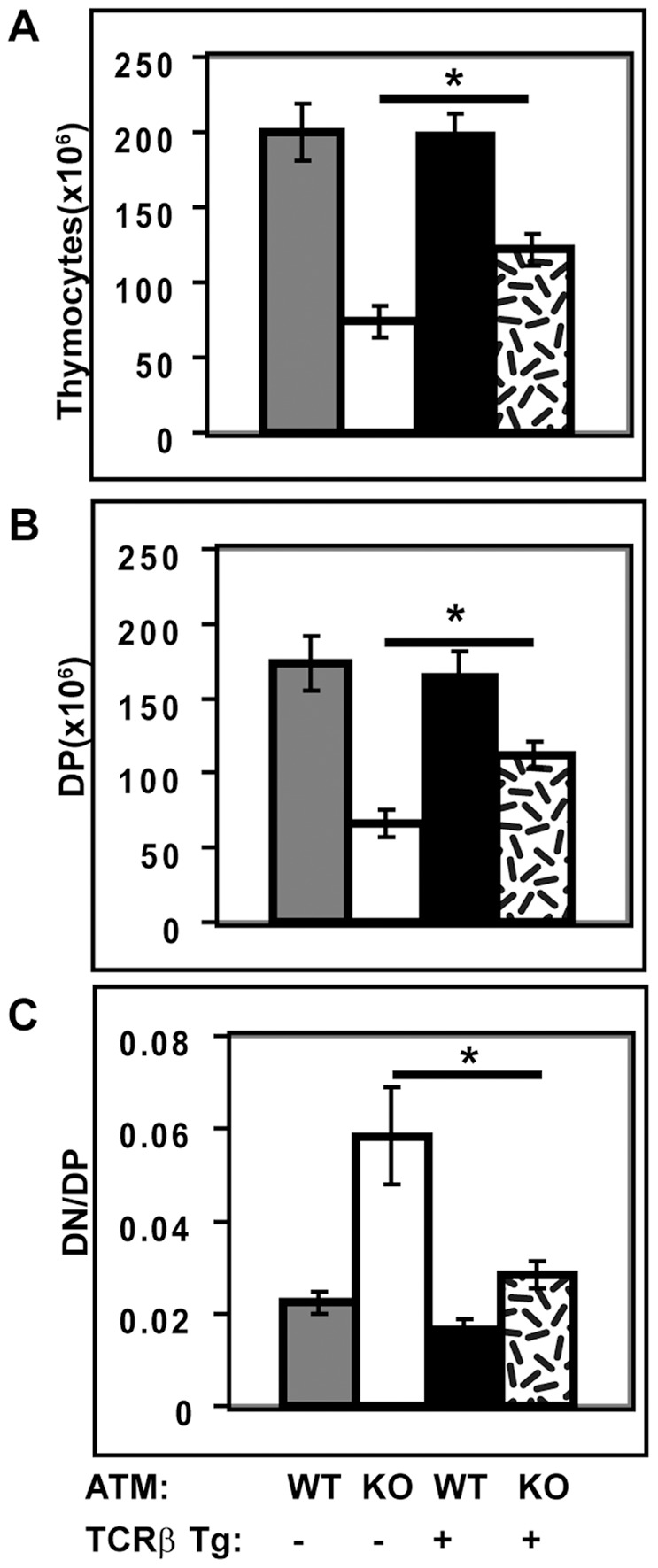
Generation and resolution of DNA double-strand breaks is required to assemble antigen-specific receptors from the genes encoding V, D, and J gene segments during recombination. The present report investigates the requirement for ataxia telangiectasia-mutated (ATM) kinase, a component of DNA double-strand break repair, during TCRβ recombination and in subsequent TCRβ-dependent repertoire generation and thymocyte development. CD4(-)CD8(-) double negative stage 2/3 thymocytes from ATM-deficient mice have both an increased frequency of cells with DNA break foci at TCRβ loci and reduced Vβ-DJβ rearrangement. Sequencing of TCRβ complementarity-determining region 3 demonstrates that ATM-deficient CD4(+)CD8(+) double positive thymocytes and peripheral T cells have altered processing of coding ends for both in-frame and out-of-frame TCRβ rearrangements, providing the unique demonstration that ATM deficiency alters the expressed TCRβ repertoire by a selection-independent mechanism. ATMKO thymi exhibit a partial developmental block in DN cells as they negotiate the β-selection checkpoint to become double negative stage 4 and CD4(+)CD8(+) thymocytes, resulting in reduced numbers of CD4(+)CD8(+) cells. Importantly, expression of a rearranged TCRβ transgene substantially reverses this defect in CD4(+)CD8(+) cells, directly linking a requirement for ATM during endogenous TCRβ rearrangement to subsequent TCRβ-dependent stages of development. These results demonstrate that ATM plays an important role in TCRβ rearrangement, generation of the TCRβ CDR3 repertoire, and efficient TCRβ-dependent T cell development.
Conflict of interest statement
Figures

References
-
- Hesslein DG, Schatz DG (2001) Factors and forces controlling V(D)J recombination. Adv Immunol 78: 169–232. - PubMed
-
- Fugmann SD, Lee AI, Shockett PE, Villey IJ, Schatz DG (2000) The RAG proteins and V(D)J recombination: complexes, ends, and transposition. Annu Rev Immunol 18: 495–527. - PubMed
-
- Goyarts EC, Vegh Z, Kalergis AM, Horig H, Papadopoulos NJ, et al. (1998) Point mutations in the beta chain CDR3 can alter the T cell receptor recognition pattern on an MHC class I/peptide complex over a broad interface area. Mol Immunol 35: 593–607. - PubMed
-
- Godfrey DI, Kennedy J, Mombaerts P, Tonegawa S, Zlotnik A (1994) Onset of TCR-beta gene rearrangement and role of TCR-beta expression during CD3–CD4–CD8- thymocyte differentiation. J Immunol 152: 4783–4792. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
