

Male and female Plasmodium falciparum mature gametocytes show different responses to antimalarial drugs
- PMID: 23629698
- PMCID: PMC3697345
- DOI: 10.1128/AAC.00325-13
Male and female Plasmodium falciparum mature gametocytes show different responses to antimalarial drugs
Abstract
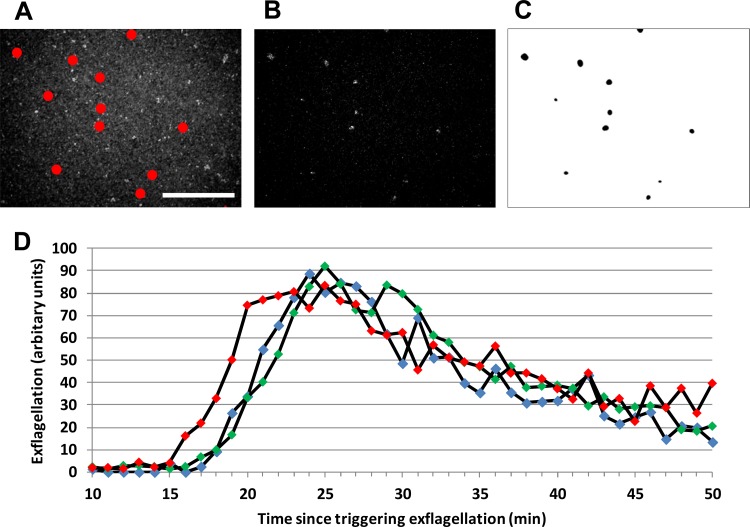
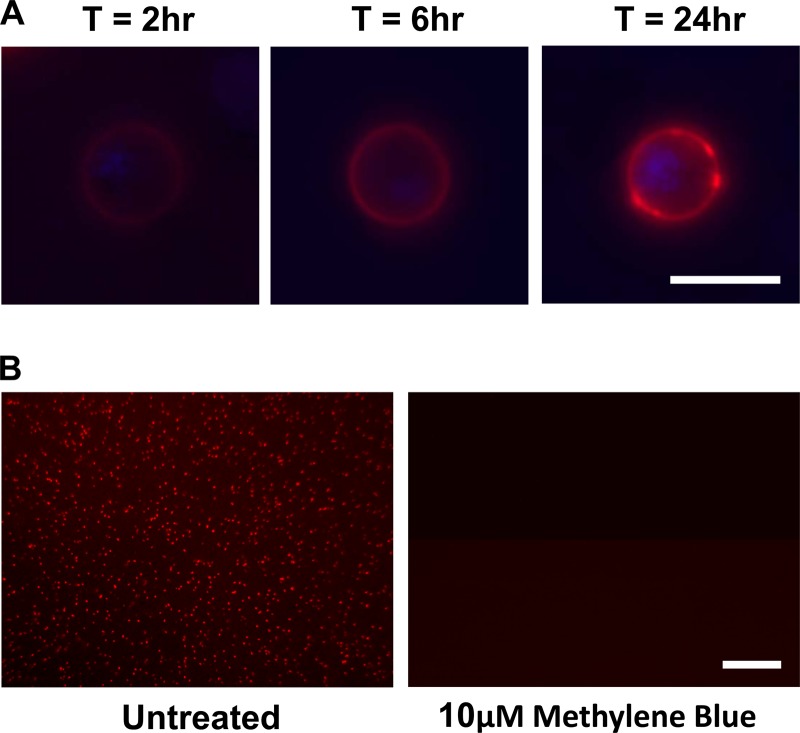
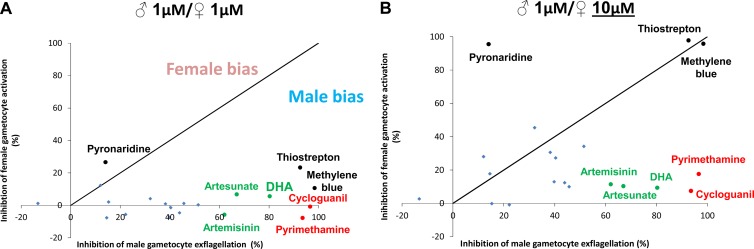
It is the mature gametocytes of Plasmodium that are solely responsible for parasite transmission from the mammalian host to the mosquito. They are therefore a logical target for transmission-blocking antimalarial interventions, which aim to break the cycle of reinfection and reduce the prevalence of malaria cases. Gametocytes, however, are not a homogeneous cell population. They are sexually dimorphic, and both males and females are required for parasite transmission. Using two bioassays, we explored the effects of 20 antimalarials on the functional viability of both male and female mature gametocytes of Plasmodium falciparum. We show that mature male gametocytes (as reported by their ability to produce male gametes, i.e., to exflagellate) are sensitive to antifolates, some endoperoxides, methylene blue, and thiostrepton, with submicromolar 50% inhibitory concentrations (IC50s), whereas female gametocytes (as reported by their ability to activate and form gametes expressing the marker Pfs25) are much less sensitive to antimalarial intervention, with only methylene blue and thiostrepton showing any significant activity. These findings show firstly that the antimalarial responses of male and female gametocytes differ and secondly that the mature male gametocyte should be considered a more vulnerable target than the female gametocyte for transmission-blocking drugs. Given the female-biased sex ratio of Plasmodium falciparum (∼3 to 5 females:1 male), current gametocyte assays without a sex-specific readout are unlikely to identify male-targeted compounds and prioritize them for further development. Both assays reported here are being scaled up to at least medium throughput and will permit identification of key transmission-blocking molecules that have been overlooked by other screening campaigns.
Figures
References
-
- World Health Organization 2011. World malaria report 2011. WHO, Geneva, Switzerland: http://www.who.int/malaria/world_malaria_report_2011/9789241564403_eng.pdf
-
- Alonso PL, Brown G, Arevalo-Herrera M, Binka F, Chitnis C, Collins F, Doumbo OK, Greenwood B, Hall BF, Levine MM, Mendis K, Newman RD, Plowe CV, Rodríguez MH, Sinden R, Slutsker L, Tanner M. 2011. A research agenda to underpin malaria eradication. PLoS Med. 8:e1000406. 10.1371/journal.pmed.1000406 - DOI - PMC - PubMed
-
- Delves M, Plouffe D, Scheurer C, Meister S, Wittlin S, Winzeler EA, Sinden RE, Leroy D. 2012. The activities of current antimalarial drugs on the life cycle stages of Plasmodium: a comparative study with human and rodent parasites. PLoS Med. 9:e1001169. 10.1371/journal.pmed.1001169 - DOI - PMC - PubMed
-
- Delves MJ, Ramakrishnan C, Blagborough AM, Leroy D, Wells TNC, Sinden RE. 2012. A high-throughput assay for the identification of malarial transmission-blocking drugs and vaccines. Int. J. Parasitol. 42:999–1006 - PubMed
-
- Lelièvre J, Almela MJ, Lozano S, Miguel C, Franco V, Leroy D, Herreros E. 2012. Activity of clinically relevant antimalarial drugs on Plasmodium falciparum mature gametocytes in an ATP bioluminescence “transmission blocking” assay. PLoS One 7:e35019. 10.1371/journal.pone.0035019 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
