

A novel approach to generate a recombinant toxoid vaccine against Clostridium difficile
- PMID: 23629868
- PMCID: PMC3749728
- DOI: 10.1099/mic.0.066712-0
A novel approach to generate a recombinant toxoid vaccine against Clostridium difficile
Abstract
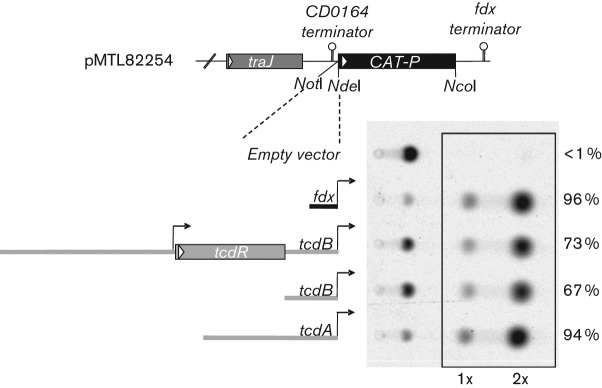
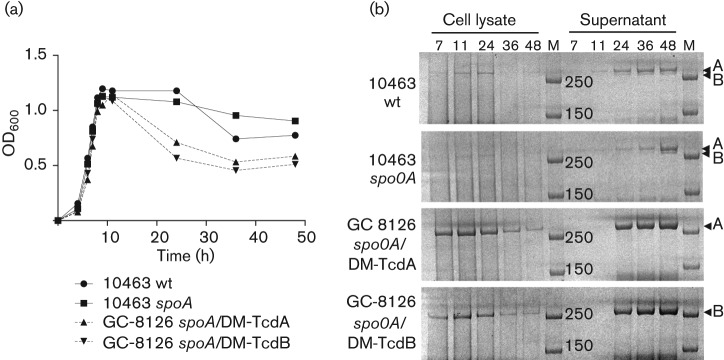
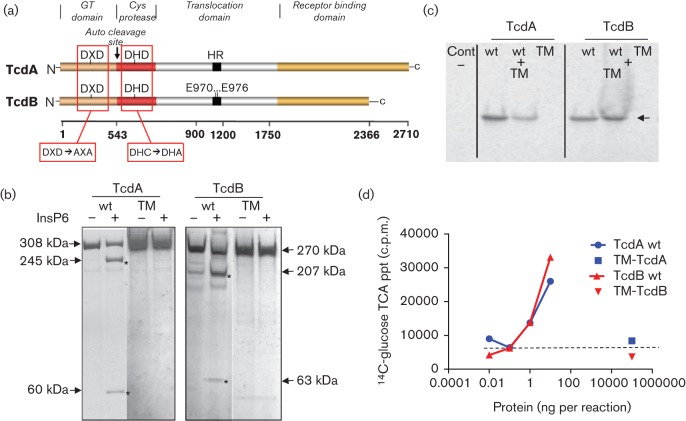
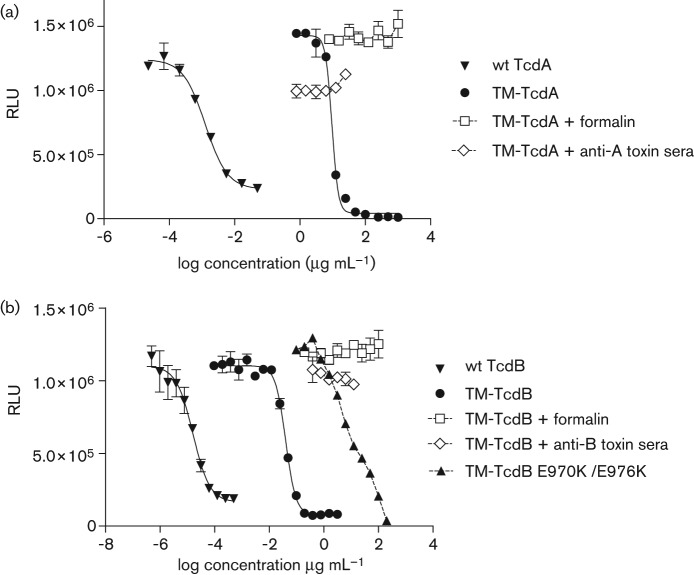
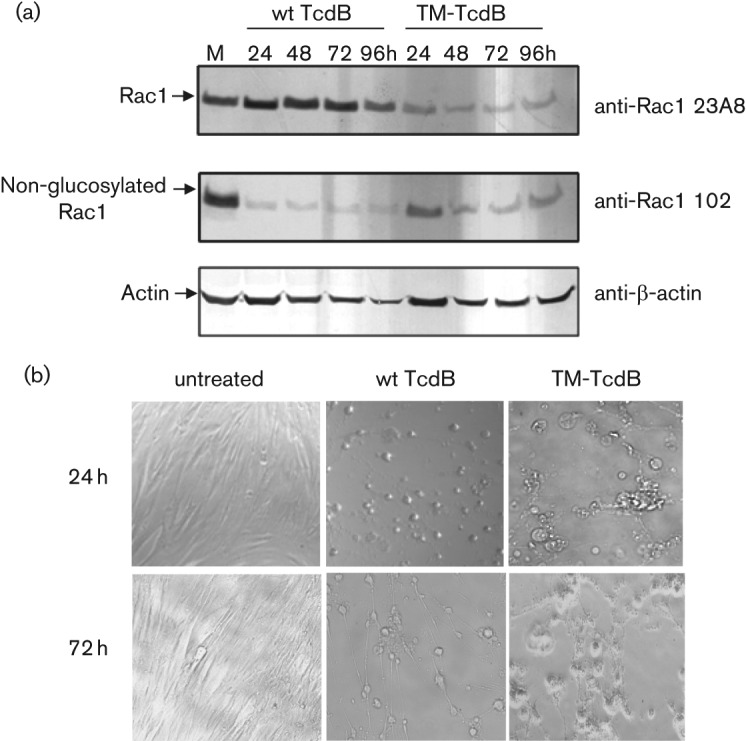
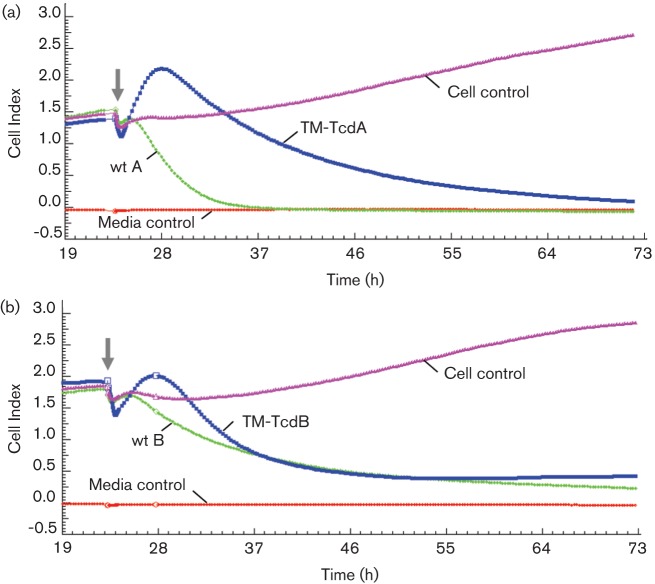
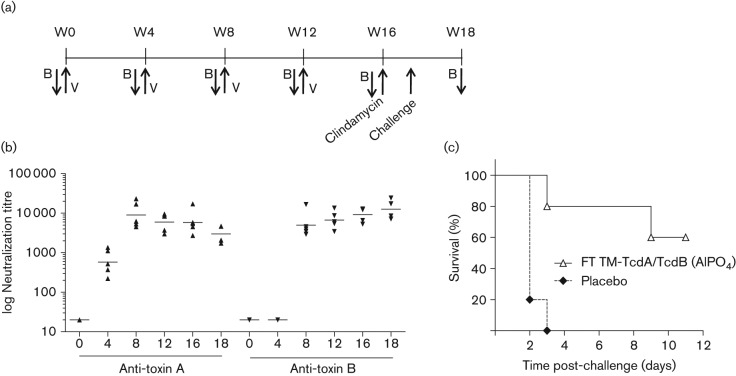
The Clostridium difficile toxins A and B are primarily responsible for symptoms of C. difficile associated disease and are prime targets for vaccine development. We describe a plasmid-based system for the production of genetically modified toxins in a non-sporulating strain of C. difficile that lacks the toxin genes tcdA and tcdB. TcdA and TcdB mutations targeting established glucosyltransferase cytotoxicity determinants were introduced into recombinant plasmids and episomally expressed toxin mutants purified from C. difficile transformants. TcdA and TcdB mutants lacking glucosyltransferase and autoproteolytic processing activities were ~10 000-fold less toxic to cultured human IMR-90 cells than corresponding recombinant or native toxins. However, both mutants retained residual cytotoxicity that could be prevented by preincubating the antigens with specific antibodies or by formalin treatment. Such non-toxic formalin-treated mutant antigens were immunogenic and protective in a hamster model of infection. The remaining toxicity of untreated TcdA and TcdB mutant antigens was associated with cellular swelling, a phenotype consistent with pore-induced membrane leakage. TcdB substitution mutations previously shown to block vesicular pore formation and toxin translocation substantially reduced residual toxicity. We discuss the implications of these results for the development of a C. difficile toxoid vaccine.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases