

Abnormal interneuron development in disrupted-in-schizophrenia-1 L100P mutant mice
- PMID: 23631734
- PMCID: PMC3648430
- DOI: 10.1186/1756-6606-6-20
Abnormal interneuron development in disrupted-in-schizophrenia-1 L100P mutant mice
Abstract
Background: Interneuron deficits are one of the most consistent findings in post-mortem studies of schizophrenia patients and are likely important in the cognitive deficits associated with schizophrenia. Disrupted-in-Schizophrenia 1 (DISC1), a strong susceptibility gene for schizophrenia and other mental illnesses, is involved in neurodevelopment, including that of interneurons. However, the mechanism by which DISC1 regulates interneuron development remains unknown. In this study, we analyzed interneuron histology in the Disc1-L100P single point mutation mouse, that was previously shown to have behavioral abnormalities and cortical developmental defects related to schizophrenia.
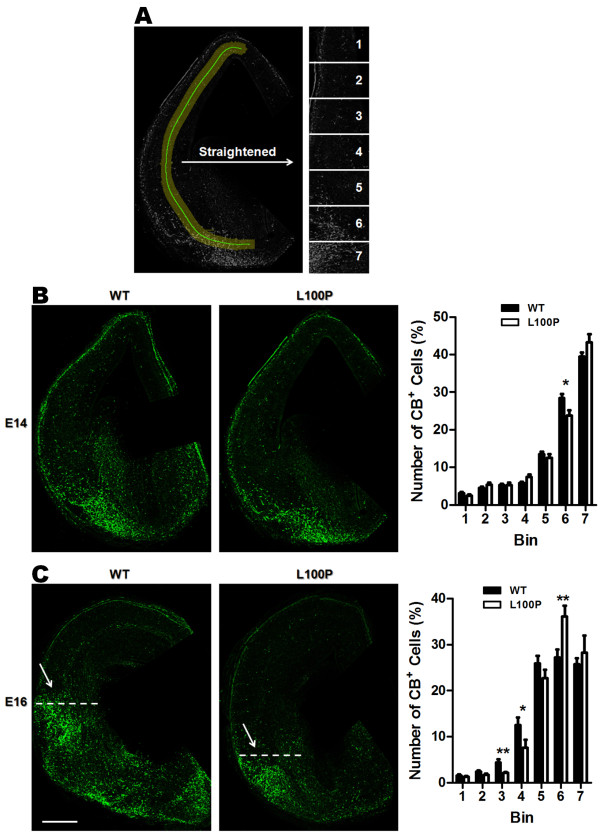
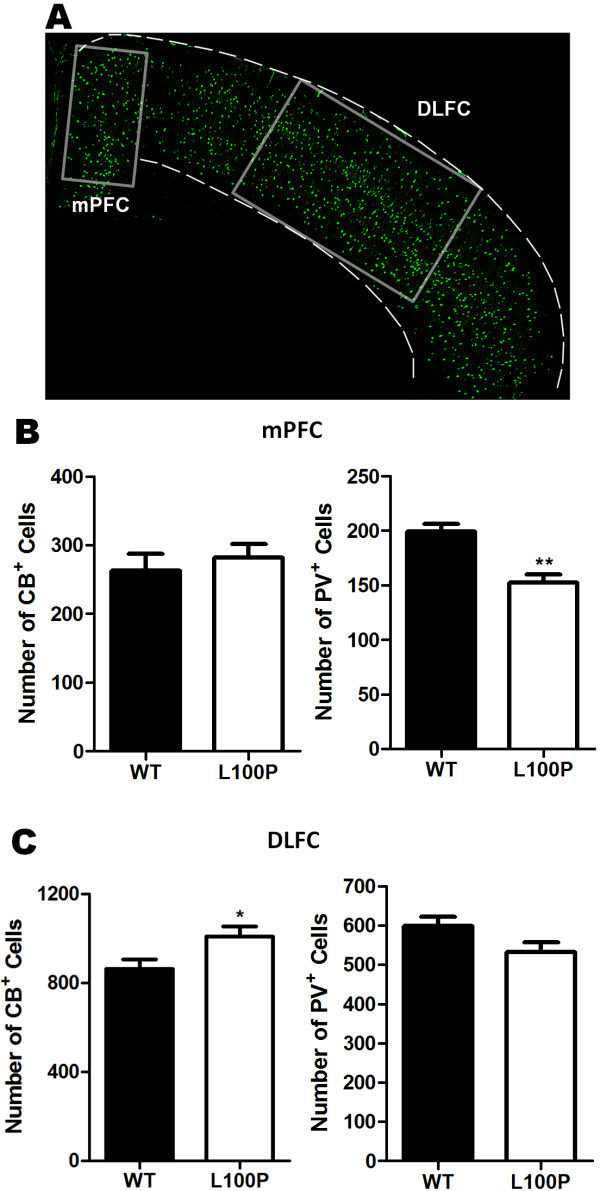
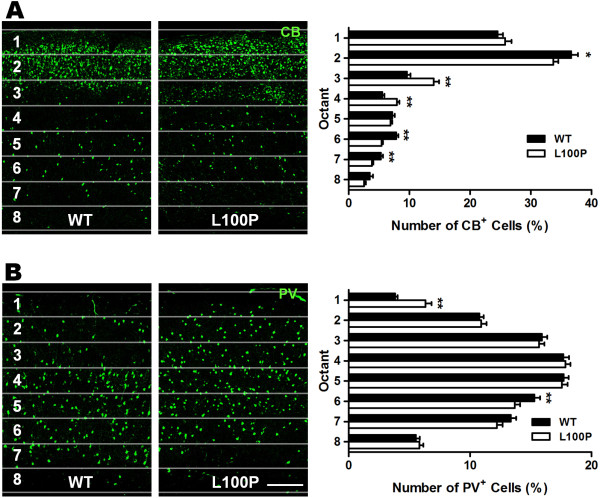
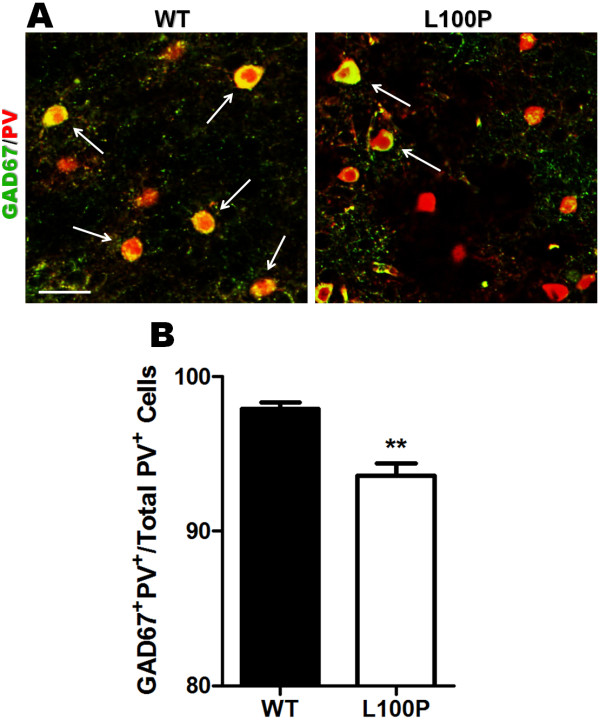
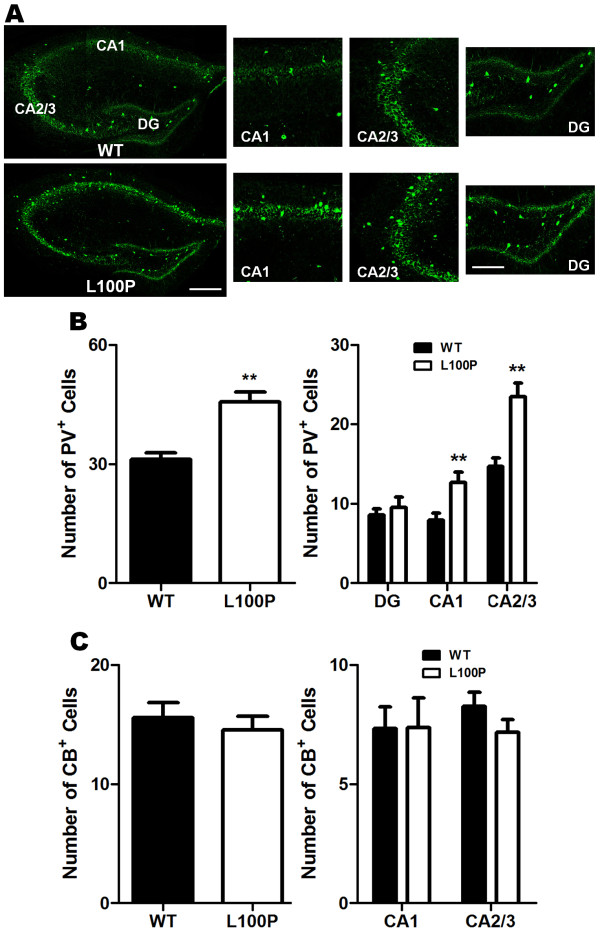
Results: We sought to determine whether a Disc1-L100P point mutation in the mouse would alter interneuron density and location. First, we examined interneuron position in the developing mouse cortex during embryonic days 14-16 as an indicator of interneuron tangential migration, and found striking migration deficits in Disc1-L100P mutants. Further analysis of adult brains revealed that the Disc1-L100P mutants have selective alterations of calbindin- and parvalbumin-expressing interneurons in the cortex and hippocampus, decreased GAD67/PV co-localization and mis-positioned interneurons across the neocortex when compared to wild-type littermates.
Conclusion: Our results are consistent with the anomalies seen in post-mortem schizophrenia studies and other Disc1 mutant mouse models. Future research is required to determine the specific mechanisms underlying these cellular deficits. Overall, these findings provide further evidence that DISC1 participates in interneuron development and add to our understanding of how DISC1 variants can affect susceptibility to psychiatric illness.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
