

STING-dependent recognition of cyclic di-AMP mediates type I interferon responses during Chlamydia trachomatis infection
- PMID: 23631912
- PMCID: PMC3663186
- DOI: 10.1128/mBio.00018-13
STING-dependent recognition of cyclic di-AMP mediates type I interferon responses during Chlamydia trachomatis infection
Abstract
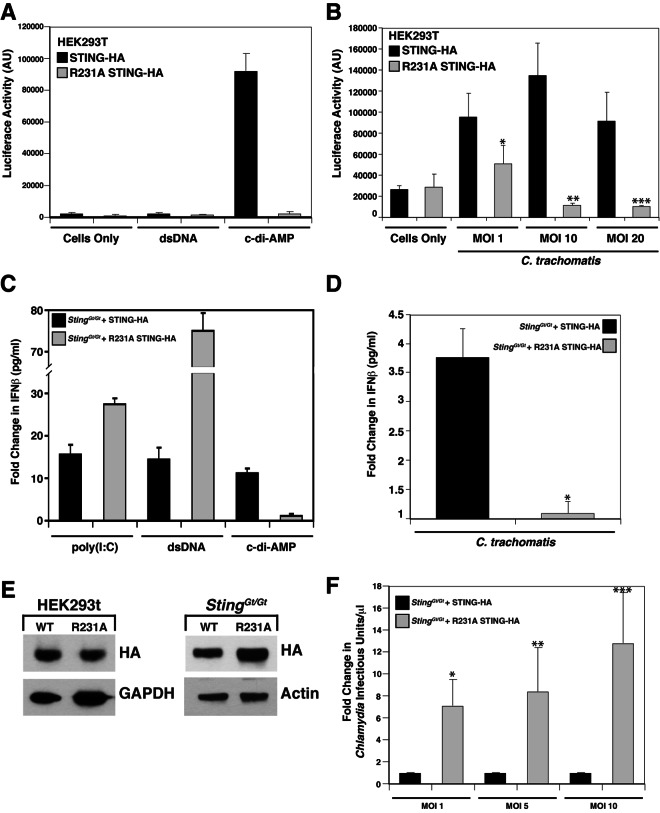
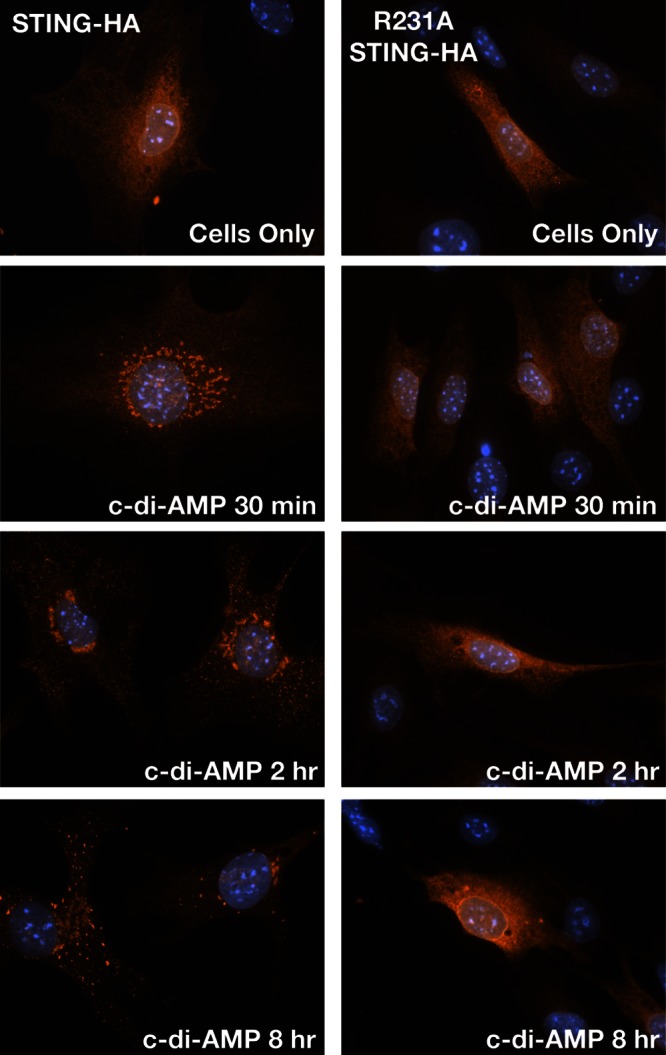
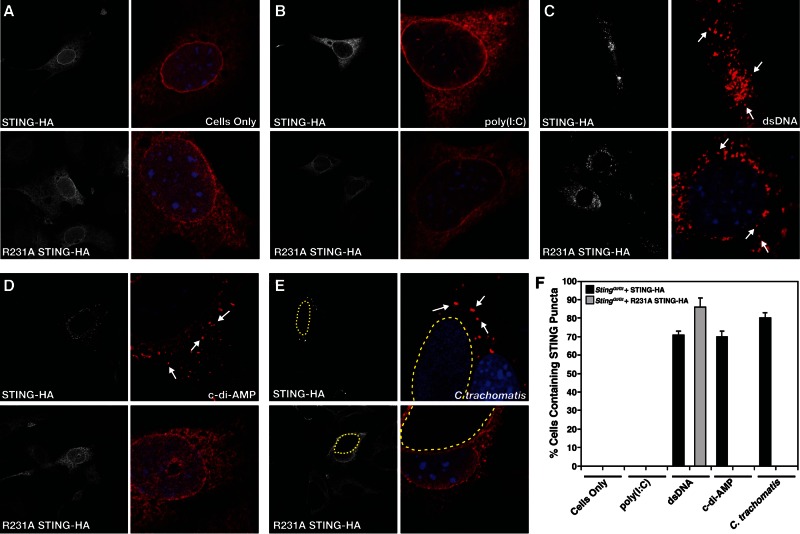
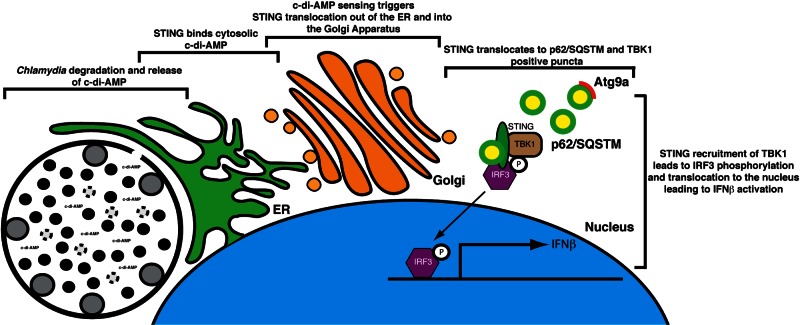
STING (stimulator of interferon [IFN] genes) initiates type I IFN responses in mammalian cells through the detection of microbial nucleic acids. The membrane-bound obligate intracellular bacterium Chlamydia trachomatis induces a STING-dependent type I IFN response in infected cells, yet the IFN-inducing ligand remains unknown. In this report, we provide evidence that Chlamydia synthesizes cyclic di-AMP (c-di-AMP), a nucleic acid metabolite not previously identified in Gram-negative bacteria, and that this metabolite is a prominent ligand for STING-mediated activation of IFN responses during infection. We used primary mouse lung fibroblasts and HEK293T cells to compare IFN-β responses to Chlamydia infection, c-di-AMP, and other type I IFN-inducing stimuli. Chlamydia infection and c-di-AMP treatment induced type I IFN responses in cells expressing STING but not in cells expressing STING variants that cannot sense cyclic dinucleotides but still respond to cytoplasmic DNA. The failure to induce a type I IFN response to Chlamydia and c-di-AMP correlated with the inability of STING to relocalize from the endoplasmic reticulum to cytoplasmic punctate signaling complexes required for IFN activation. We conclude that Chlamydia induces STING-mediated IFN responses through the detection of c-di-AMP in the host cell cytosol and propose that c-di-AMP is the ligand predominantly responsible for inducing such a response in Chlamydia-infected cells.
Importance: This study shows that the Gram-negative obligate pathogen Chlamydia trachomatis, a major cause of pelvic inflammatory disease and infertility, synthesizes cyclic di-AMP (c-di-AMP), a nucleic acid metabolite that thus far has been described only in Gram-positive bacteria. We further provide evidence that the host cell employs an endoplasmic reticulum (ER)-localized cytoplasmic sensor, STING (stimulator of interferon [IFN] genes), to detect c-di-AMP synthesized by Chlamydia and induce a protective IFN response. This detection occurs even though Chlamydia is confined to a membrane-bound vacuole. This raises the possibility that the ER, an organelle that innervates the entire cytoplasm, is equipped with pattern recognition receptors that can directly survey membrane-bound pathogen-containing vacuoles for leaking microbe-specific metabolites to mount type I IFN responses required to control microbial infections.
Figures
References
-
- Haggerty CL, Gottlieb SL, Taylor BD, Low N, Xu F, Ness RB. 2010. Risk of sequelae after Chlamydia trachomatis genital infection in women. J. Infect. Dis. 201:S134–S155 - PubMed
-
- Abdelrahman YM, Belland RJ. 2005. The chlamydial developmental cycle. FEMS Microbiol. Rev. 29:949–959 - PubMed
-
- Takeuchi O, Akira S. 2010. Pattern recognition receptors and inflammation. Cell 140:805–820 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R21 AI103817/AI/NIAID NIH HHS/United States
- F32 AI096866/AI/NIAID NIH HHS/United States
- AI080749/AI/NIAID NIH HHS/United States
- 2-U54-AI-057153/AI/NIAID NIH HHS/United States
- 1R01-AI081694/AI/NIAID NIH HHS/United States
- AI063302/AI/NIAID NIH HHS/United States
- P01 AI063302/AI/NIAID NIH HHS/United States
- R01 AI081694/AI/NIAID NIH HHS/United States
- U54 AI057153/AI/NIAID NIH HHS/United States
- R01 AI080749/AI/NIAID NIH HHS/United States
- T32 GM007184/GM/NIGMS NIH HHS/United States
- AI096866-02/AI/NIAID NIH HHS/United States
- F32 AI091100/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
