

Review
doi: 10.1074/jbc.R113.452805.
Epub 2013 Apr 30.
Structural diversity of eukaryotic membrane cytochrome p450s
Affiliations
- PMID: 23632020
- PMCID: PMC3682514
- DOI: 10.1074/jbc.R113.452805
Item in Clipboard
Review
Structural diversity of eukaryotic membrane cytochrome p450s
J Biol Chem.
.
Abstract
X-ray crystal structures are available for 29 eukaryotic microsomal, chloroplast, or mitochondrial cytochrome P450s, including two non-monooxygenase P450s. These structures provide a basis for understanding structure-function relations that underlie their distinct catalytic activities. Moreover, structural plasticity has been characterized for individual P450s that aids in understanding substrate binding in P450s that mediate drug clearance.
Keywords: Carcinogenesis; Cytochrome P450; Drug Development; Drug Metabolism; Protein structure; Steroidogenesis.
Figures
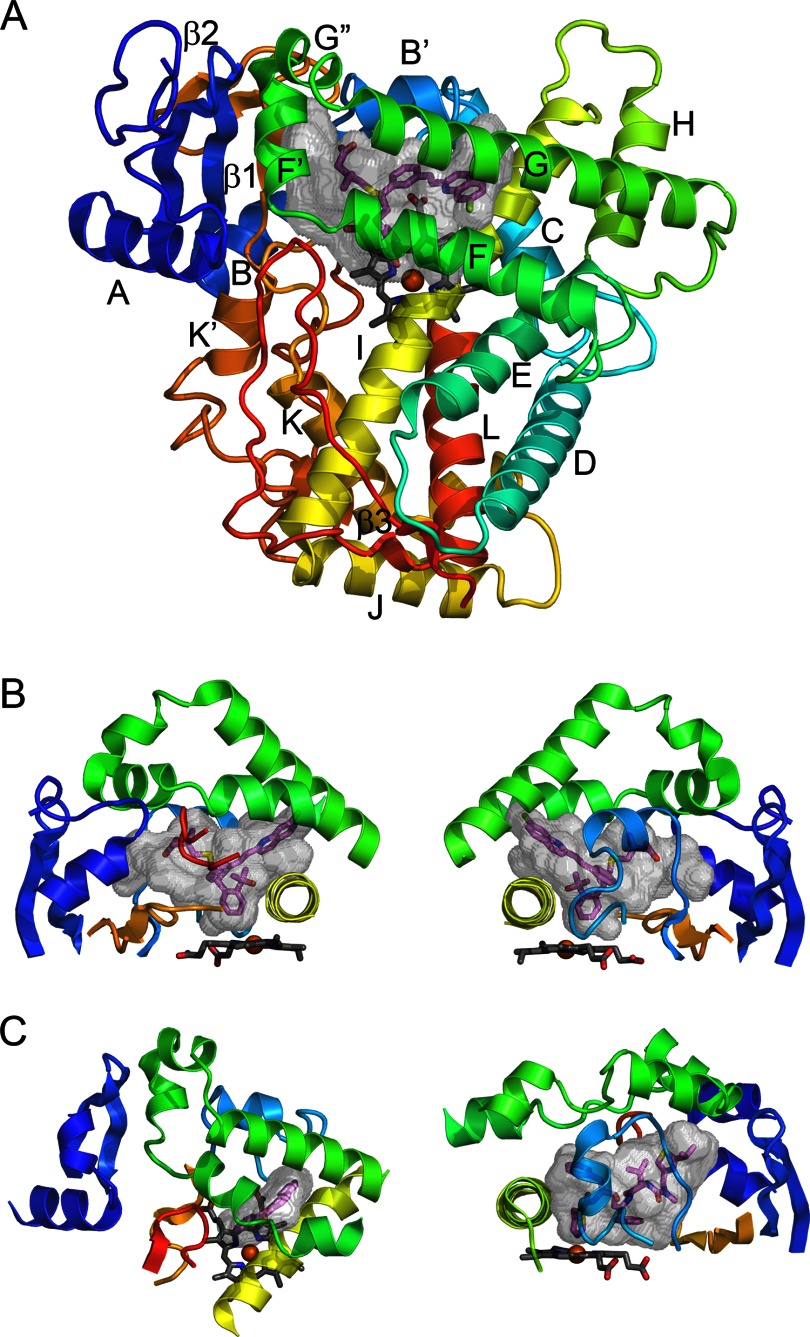
P450 fold and elements comprising the active site.
A, the overall topology is illustrated by the structure of microsomal 2C8 (Protein Data Bank code 2NNI), colored from blue at the N terminus to red at the C terminus. The active site cavity is shown as a transparent surface. The bound substrate, montelukast (violet carbons), and the heme prosthetic group (gray carbons) are shown as stick figures. Twelve helices designated by letters and β-sheets 1 and 2 are highly conserved. Additional helices that are named by letters with primes or double primes are evident. B, two views of structural components that form the sides of the substrate-binding site of 2C8. The helix F-G region (green) forms the top of the cavity and is cantilevered over helix I (yellow), which forms one side. The opposite side is formed by connections (orange) between helix K and β-strands 1–3 and between β-strands 1–4 and helix K′ near the surface of the heme and by the N-terminal region (dark blue) that includes helix A and β-strand 1. The gaps under the helix F-G region between helix I and the N-terminal region are filled by the C-terminal loop (red orange) as shown in the left panel and by the B-C loop (light blue) as shown in the right panel. C, views of the helix F-G side of the microsomal 1A2 complex with α-naphthoflavone (left) and of the B-C loop side of the microsomal 3A4 complex with ritonavir (right) illustrate differences in the topologies of the active sites and the secondary and tertiary structures of 2C8 (code 2NNI), 1A2 (code 2HI4), and 3A4 (code 3NXU).
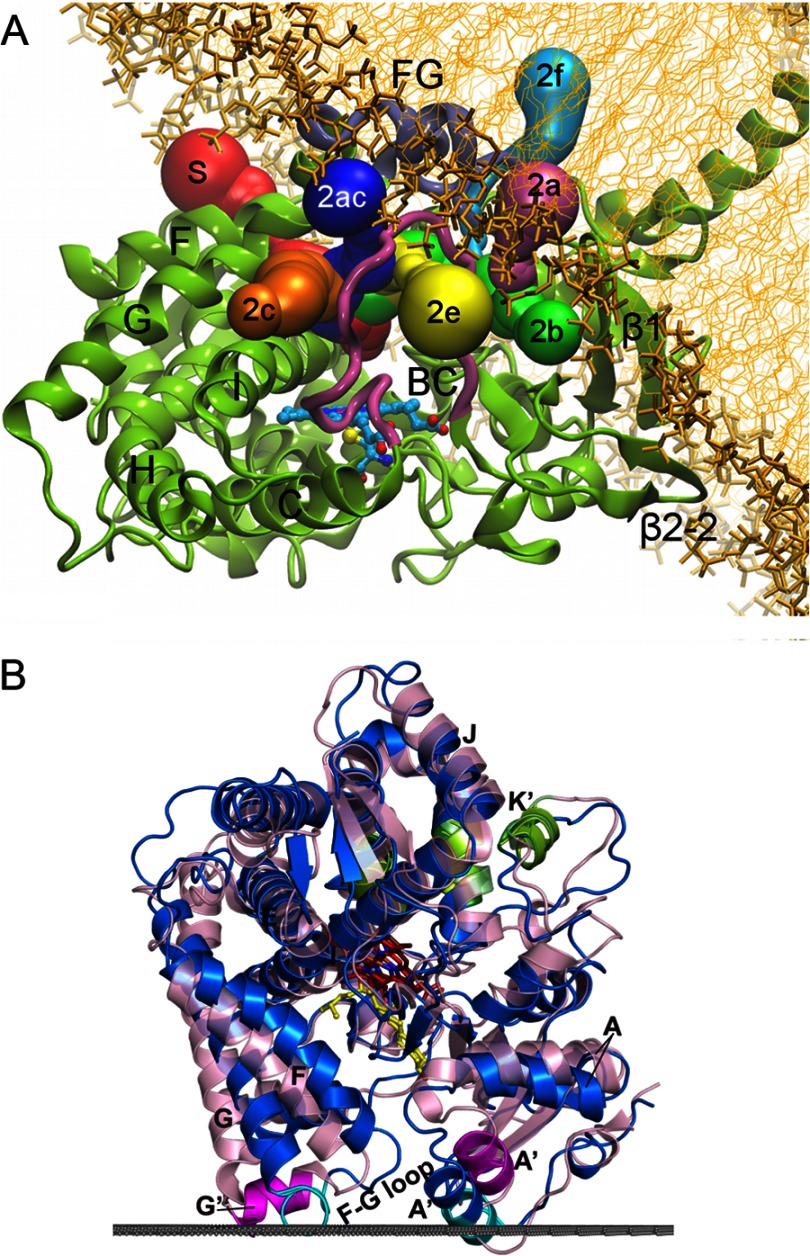
Interactions of microsomal (A) and mitochondrial (B) P450s with the membrane.
A, Fig. 5A from Cojocaru et al. (32) reproduced here under the terms of the Creative Commons Attribution License. A model of microsomal 2C9 (ribbon) embedded in a 1-palmitoyl-2-oleoylphosphatidylcholine bilayer (thick and thin gold sticks) is depicted. The colored tubes represent solvent access channels that were observed to open and, in some cases, close during molecular dynamics simulations and are labeled according to the nomenclature of Cojocaru et al. (19). The region between helices F and G is designated FG. B, Fig. S6 from Mast et al. (33). Structures of 11A1 (blue; Protein Data Bank code 3MZS) and 24A1 (pink; code 3K9V) are shown. The upper surface of the membrane bilayer with respect to 11A1 is indicated by the gray line. The membrane insertion sequences are colored cyan in 11A1 and magenta in 24A1. Secondary structural elements that participate in adrenodoxin binding are colored green. 22-Hydroxycholesterol is shown in yellow; heme is shown in red in 11A1 and brown in 24A1.
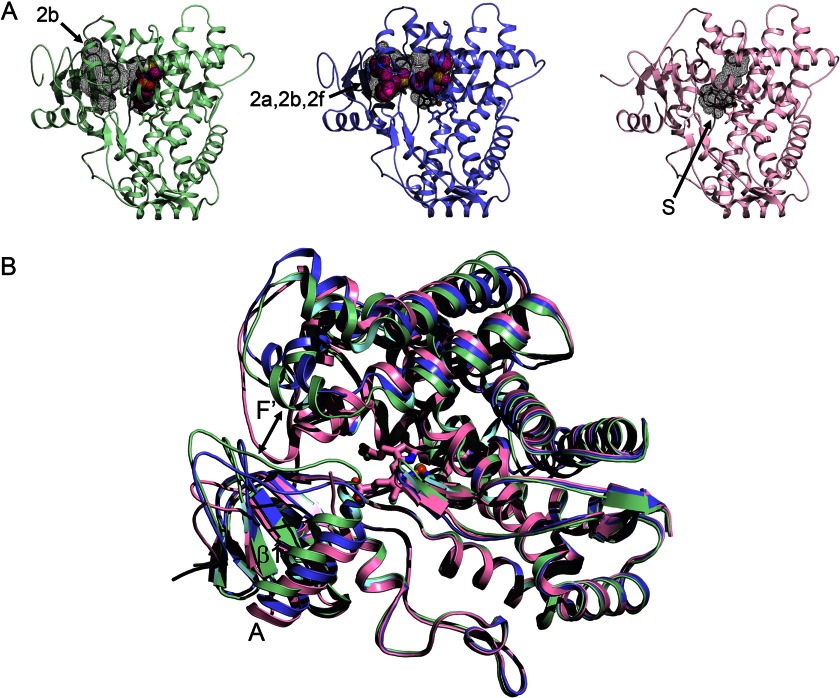
Alternative conformations of microsomal 2D6.
A, the 3QM4 structure of 2D6 with prinomastat bound (green) exhibits a closed active site cavity and an antechamber (black mesh surfaces) with an open 2b channel (arrow). The 3TBG structure of 2D6 with two molecules of thioridazine bound (slate) displays a single open cavity with merged 2a, 2b, and 2f channels. The 2F9Q structure of 2D6 (pink) exhibits an open S channel. B, a side view of the overlaid structures shows how conformational changes open the entrance channel in the 3TBG structure relative to the 3QM4 structure (double-headed arrow), whereas the F-G loop fills the channel entrance in the 2F9Q structure.
References
-
- Nelson D. R., Zeldin D. C., Hoffman S. M., Maltais L. J., Wain H. M., Nebert D. W. (2004) Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics 14, 1–18 - PubMed
-
- Williams P. A., Cosme J., Sridhar V., Johnson E. F., McRee D. E. (2000) The crystallographic structure of a mammalian microsomal cytochrome P450 monooxygenase: structural adaptations for membrane binding and functional diversity. Mol. Cell 5, 121–131 - PubMed
-
- Poulos T. L., Finzel B. C., Gunsalus I. C., Wagner G. C., Kraut J. (1985) The 2.6-Å crystal structure of the Pseudomonas putida cytochrome P-450. J. Biol. Chem. 260, 16122–16130 - PubMed
-
- Poulos T. L., Johnson E. F. (2005) Structures of cytochrome P450 enzymes. In Cytochrome P450: Structure, Mechanism, and Biochemistry (Ortiz de Montellano P. R., ed) 3rd Ed., pp. 87–114, Kluwer Academic/Plenum Publishers, New York
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
