

Fast neurotransmitter release regulated by the endocytic scaffold intersectin
- PMID: 23633571
- PMCID: PMC3657817
- DOI: 10.1073/pnas.1219234110
Fast neurotransmitter release regulated by the endocytic scaffold intersectin
Abstract
Sustained fast neurotransmission requires the rapid replenishment of release-ready synaptic vesicles (SVs) at presynaptic active zones. Although the machineries for exocytic fusion and for subsequent endocytic membrane retrieval have been well characterized, little is known about the mechanisms underlying the rapid recruitment of SVs to release sites. Here we show that the Down syndrome-associated endocytic scaffold protein intersectin 1 is a crucial factor for the recruitment of release-ready SVs. Genetic deletion of intersectin 1 expression or acute interference with intersectin function inhibited the replenishment of release-ready vesicles, resulting in short-term depression, without significantly affecting the rate of endocytic membrane retrieval. Acute perturbation experiments suggest that intersectin-mediated vesicle replenishment involves the association of intersectin with the fissioning enzyme dynamin and with the actin regulatory GTPase CDC42. Our data indicate a role for the endocytic scaffold intersectin in fast neurotransmitter release, which may be of prime importance for information processing in the brain.
Keywords: endocytosis; synaptic transmission; synaptic vesicle recruitment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
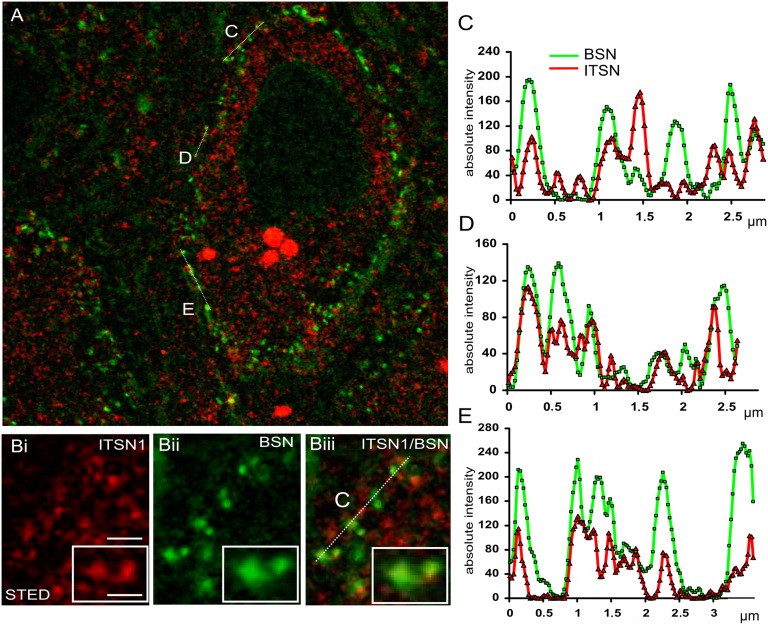
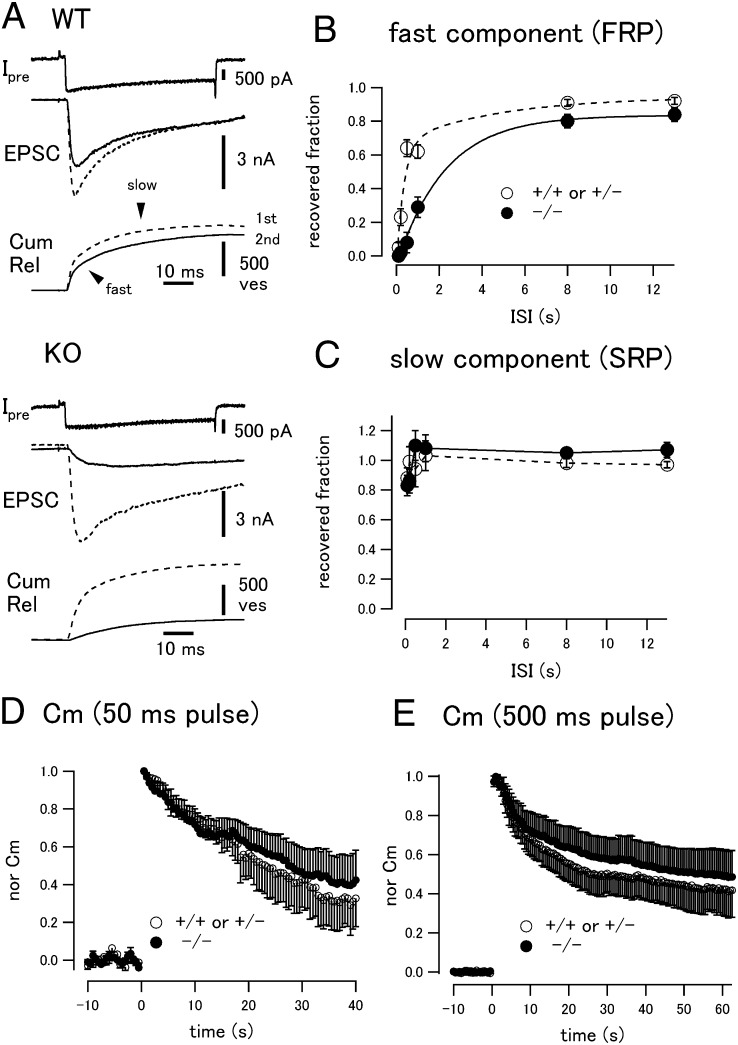
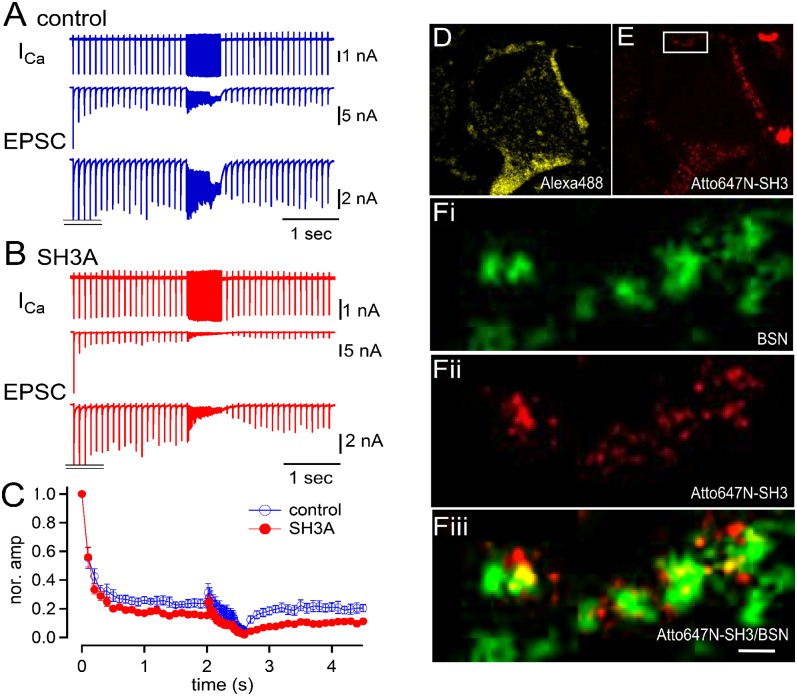


Similar articles
-
Intersectin associates with synapsin and regulates its nanoscale localization and function.Proc Natl Acad Sci U S A. 2017 Nov 7;114(45):12057-12062. doi: 10.1073/pnas.1715341114. Epub 2017 Oct 23. Proc Natl Acad Sci U S A. 2017. PMID: 29078407 Free PMC article.
-
Intersectin-Mediated Clearance of SNARE Complexes Is Required for Fast Neurotransmission.Cell Rep. 2020 Jan 14;30(2):409-420.e6. doi: 10.1016/j.celrep.2019.12.035. Cell Rep. 2020. PMID: 31940485
-
Intersectin 1: a versatile actor in the synaptic vesicle cycle.Biochem Soc Trans. 2010 Feb;38(Pt 1):181-6. doi: 10.1042/BST0380181. Biochem Soc Trans. 2010. PMID: 20074056 Review.
-
Vesicle uncoating regulated by SH3-SH3 domain-mediated complex formation between endophilin and intersectin at synapses.EMBO Rep. 2015 Feb;16(2):232-9. doi: 10.15252/embr.201439260. Epub 2014 Dec 17. EMBO Rep. 2015. PMID: 25520322 Free PMC article.
-
[Function of intersectin in endocytosis and exocytosis].Sheng Li Xue Bao. 2012 Aug 25;64(4):489-94. Sheng Li Xue Bao. 2012. PMID: 22907312 Review. Chinese.
Cited by
-
Microtubule assembly by tau impairs endocytosis and neurotransmission via dynamin sequestration in Alzheimer's disease synapse model.Elife. 2022 Apr 26;11:e73542. doi: 10.7554/eLife.73542. Elife. 2022. PMID: 35471147 Free PMC article.
-
Safeguards of Neurotransmission: Endocytic Adaptors as Regulators of Synaptic Vesicle Composition and Function.Front Cell Neurosci. 2017 Oct 11;11:320. doi: 10.3389/fncel.2017.00320. eCollection 2017. Front Cell Neurosci. 2017. PMID: 29085282 Free PMC article. Review.
-
Presynaptic Rac1 controls synaptic strength through the regulation of synaptic vesicle priming.Elife. 2022 Oct 10;11:e81505. doi: 10.7554/eLife.81505. Elife. 2022. PMID: 36214784 Free PMC article.
-
Endophilin-A coordinates priming and fusion of neurosecretory vesicles via intersectin.Nat Commun. 2020 Mar 9;11(1):1266. doi: 10.1038/s41467-020-14993-8. Nat Commun. 2020. PMID: 32152276 Free PMC article.
-
Intersectin: The Crossroad between Vesicle Exocytosis and Endocytosis.Front Endocrinol (Lausanne). 2013 Aug 27;4:109. doi: 10.3389/fendo.2013.00109. eCollection 2013. Front Endocrinol (Lausanne). 2013. PMID: 23986746 Free PMC article.
References
-
- Zucker RS, Regehr WG. Short-term synaptic plasticity. Annu Rev Physiol. 2002;64:355–405. - PubMed
-
- Neher E, Sakaba T. Multiple roles of calcium ions in the regulation of neurotransmitter release. Neuron. 2008;59(6):861–872. - PubMed
-
- Stevens CF. Neurotransmitter release at central synapses. Neuron. 2003;40(2):381–388. - PubMed
-
- Südhof TC. The synaptic vesicle cycle. Annu Rev Neurosci. 2004;27:509–547. - PubMed
-
- Rosenmund C, Rettig J, Brose N. Molecular mechanisms of active zone function. Curr Opin Neurobiol. 2003;13(5):509–519. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
