

Expression and cellular trafficking of GP82 and GP90 glycoproteins during Trypanosoma cruzi metacyclogenesis
- PMID: 23634710
- PMCID: PMC3652755
- DOI: 10.1186/1756-3305-6-127
Expression and cellular trafficking of GP82 and GP90 glycoproteins during Trypanosoma cruzi metacyclogenesis
Abstract
Background: The transformation of noninfective epimastigotes into infective metacyclic trypomastigotes (metacyclogenesis) is a fundamental step in the life cycle of Trypanosoma cruzi, comprising several morphological and biochemical changes. GP82 and GP90 are glycoproteins expressed at the surface of metacyclic trypomastigote, with opposite roles in mammalian cell invasion. GP82 is an adhesin that promotes cell invasion, while GP90 acts as a negative regulator of parasite internalization. Our understanding of the synthesis and intracellular trafficking of GP82 and GP90 during metacyclogenesis is still limited. Therefore, we decided to determine whether GP82 and GP90 are expressed only in fully differentiated metacyclic forms or they start to be expressed in intermediate forms undergoing differentiation.
Methods: Parasite populations enriched in intermediate forms undergoing differentiation were analyzed by quantitative real-time PCR, Western blot, flow cytometry and immunofluorescence to assess GP82 and GP90 expression.
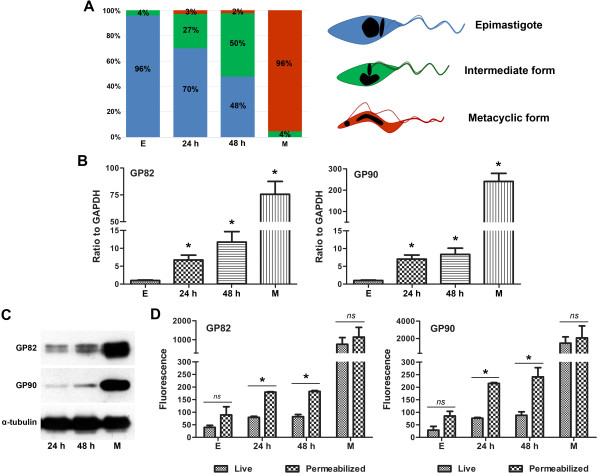
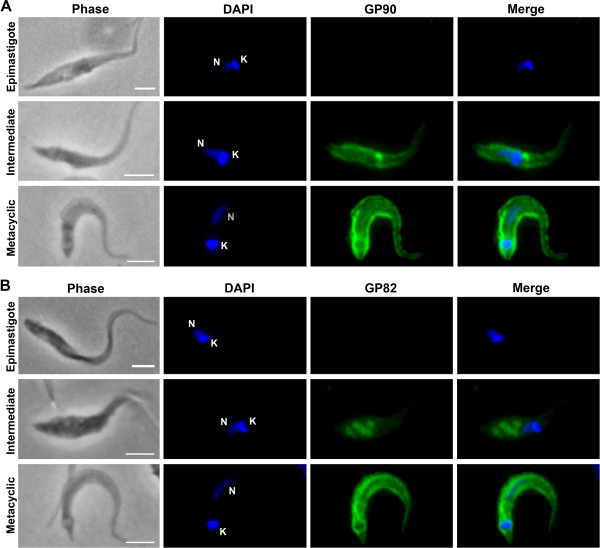
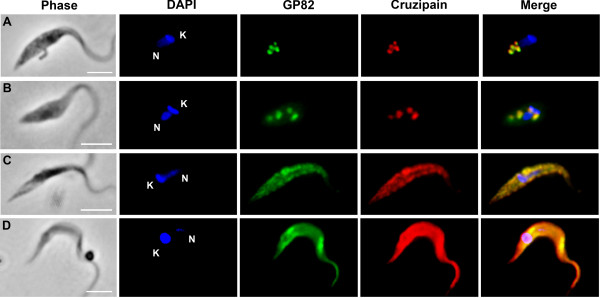
Results: We found that GP82 and GP90 mRNAs and proteins are expressed in intermediate forms and reach higher levels in fully differentiated metacyclic forms. Surprisingly, GP82 and GP90 presented distinct cellular localizations in intermediate forms compared to metacyclic trypomastigotes. In intermediate forms, GP82 is localized in organelles at the posterior region and colocalizes with cruzipain, while GP90 is localized at the flagellar pocket region.
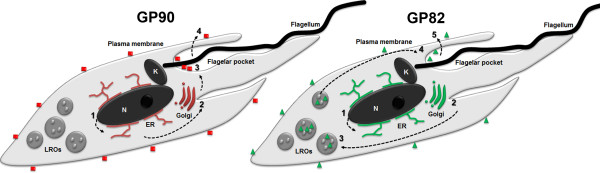
Conclusions: This study discloses new aspects of protein expression and trafficking during T. cruzi differentiation by showing that the machinery involved in GP82 and GP90 gene expression starts to operate early in the differentiation process and that different secretion pathways are responsible for delivering these glycoproteins toward the cell surface.
Figures
Similar articles
-
Expression of GP82 and GP90 surface glycoprotein genes of Trypanosoma cruzi during in vivo metacyclogenesis in the insect vector Rhodnius prolixus.Acta Trop. 2008 Jan;105(1):87-91. doi: 10.1016/j.actatropica.2007.08.004. Epub 2007 Aug 21. Acta Trop. 2008. PMID: 17889817
-
Posttranscriptional mechanisms involved in the control of expression of the stage-specific GP82 surface glycoprotein in Trypanosoma cruzi.Acta Trop. 2009 Feb;109(2):152-8. doi: 10.1016/j.actatropica.2008.10.006. Epub 2008 Nov 1. Acta Trop. 2009. PMID: 19013421
-
Shedding of Trypanosoma cruzi Surface Molecules That Regulate Host Cell Invasion Involves Phospholipase C and Increases Upon Sterol Depletion.Front Cell Infect Microbiol. 2021 Oct 19;11:769722. doi: 10.3389/fcimb.2021.769722. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34737979 Free PMC article.
-
Trypanosoma cruzi infection by oral route: how the interplay between parasite and host components modulates infectivity.Parasitol Int. 2008 Jun;57(2):105-9. doi: 10.1016/j.parint.2007.12.008. Epub 2007 Dec 23. Parasitol Int. 2008. PMID: 18234547 Review.
-
Genetic structure and expression of the surface glycoprotein GP82, the main adhesin of Trypanosoma cruzi metacyclic trypomastigotes.ScientificWorldJournal. 2013;2013:156734. doi: 10.1155/2013/156734. Epub 2013 Feb 4. ScientificWorldJournal. 2013. PMID: 23431251 Free PMC article. Review.
Cited by
-
Colchicine treatment reversibly blocks cytokinesis but not mitosis in Trypanosoma cruzi epimastigotes.Parasitol Res. 2015 Feb;114(2):641-9. doi: 10.1007/s00436-014-4227-8. Epub 2014 Nov 20. Parasitol Res. 2015. PMID: 25407128
-
All Roads Lead to Cytosol: Trypanosoma cruzi Multi-Strategic Approach to Invasion.Front Cell Infect Microbiol. 2021 Mar 5;11:634793. doi: 10.3389/fcimb.2021.634793. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 33747982 Free PMC article. Review.
-
The RNA-binding protein TcUBP1 up-regulates an RNA regulon for a cell surface-associated Trypanosoma cruzi glycoprotein and promotes parasite infectivity.J Biol Chem. 2019 Jun 28;294(26):10349-10364. doi: 10.1074/jbc.RA118.007123. Epub 2019 May 21. J Biol Chem. 2019. PMID: 31113862 Free PMC article.
-
Transcriptional remodeling during metacyclogenesis in Trypanosoma cruzi I.Virulence. 2020 Dec;11(1):969-980. doi: 10.1080/21505594.2020.1797274. Virulence. 2020. PMID: 32715914 Free PMC article.
-
Transcriptomic changes across the life cycle of Trypanosoma cruzi II.PeerJ. 2020 May 14;8:e8947. doi: 10.7717/peerj.8947. eCollection 2020. PeerJ. 2020. PMID: 32461822 Free PMC article.
References
-
- de Souza W. Cell biology of Trypanosoma cruzi. Int Rev Cytol. 1984;86:197–283. - PubMed
-
- Contreras VT, Araujo-Jorge TC, Bonaldo MC, Thomaz N, Barbosa HS, Meirelles Mde N, Goldenberg S. Biological aspects of the Dm 28c clone of Trypanosoma cruzi after metacyclogenesis in chemically defined media. Mem Inst Oswaldo Cruz. 1988;83(1):123–133. doi: 10.1590/S0074-02761988000100016. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
