

Product feedback regulation implicated in translational control of the Trypanosoma brucei S-adenosylmethionine decarboxylase regulatory subunit prozyme
- PMID: 23634831
- PMCID: PMC3771639
- DOI: 10.1111/mmi.12226
Product feedback regulation implicated in translational control of the Trypanosoma brucei S-adenosylmethionine decarboxylase regulatory subunit prozyme
Abstract
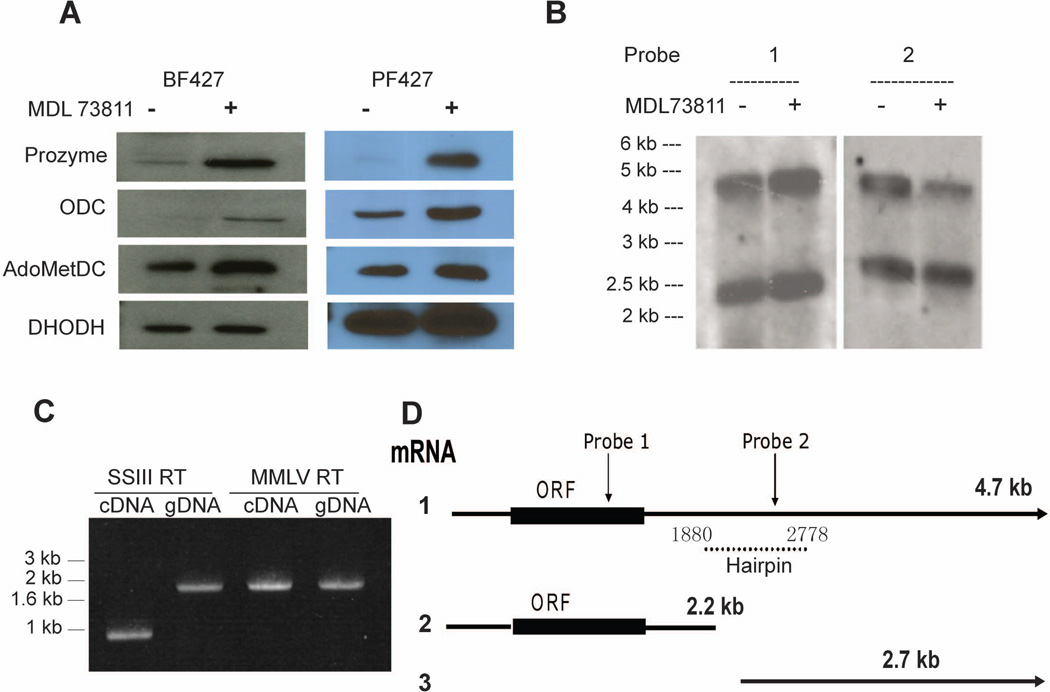
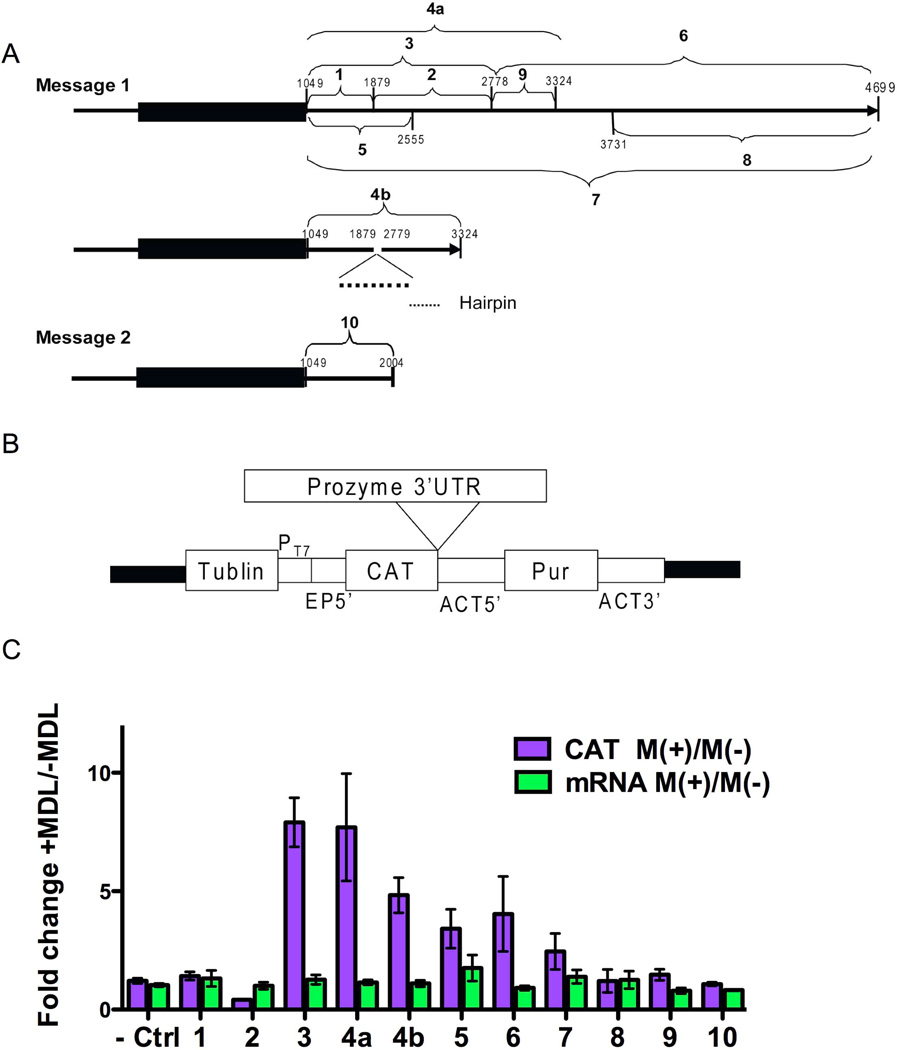
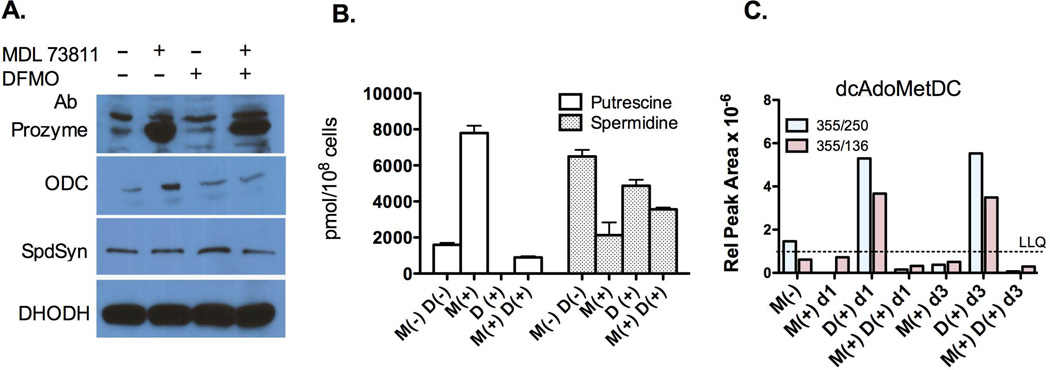
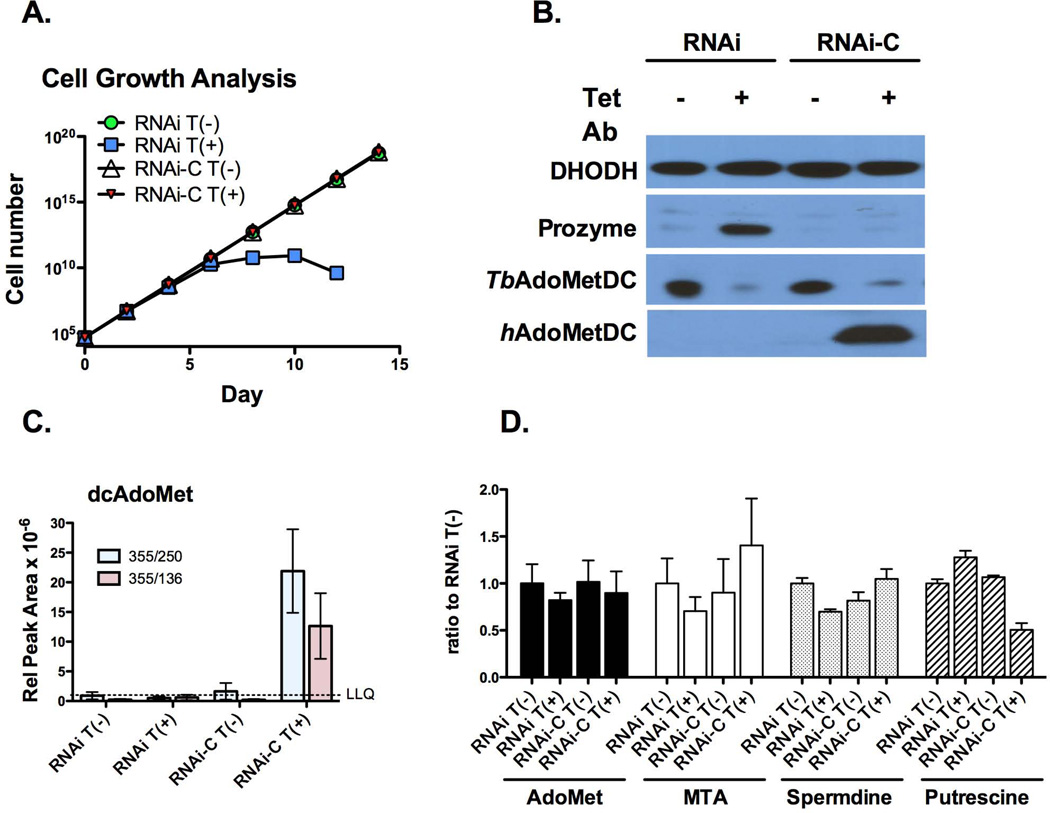
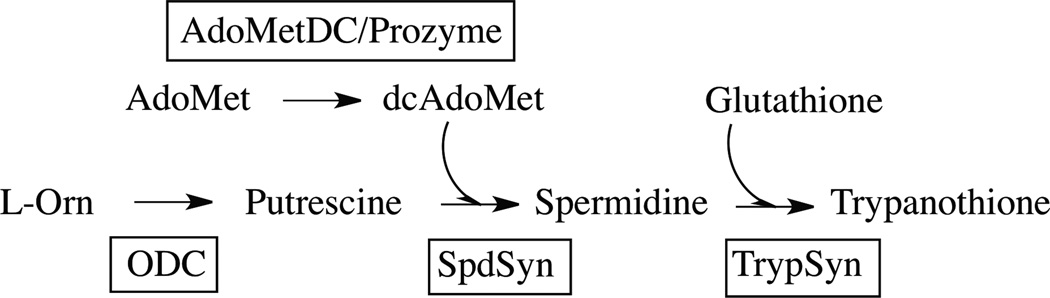
Human African sleeping sickness (HAT) is caused by the parasitic protozoan Trypanosoma brucei. Polyamine biosynthesis is an important drug target in the treatment of HAT. Previously we showed that trypanosomatid S-adenosylmethionine decarboxylase (AdoMetDC), a key enzyme for biosynthesis of the polyamine spermidine, is activated by heterodimer formation with an inactive paralogue termed prozyme. Furthermore, prozyme protein levels were regulated in response to reduced AdoMetDC activity. Herein we show that T. brucei encodes three prozyme transcripts. The 3'UTRs of these transcripts were mapped and chloramphenicol acetyltransferase (CAT) reporter constructs were used to identify a 1.2 kb region that contained a 3'UTR prozyme regulatory element sufficient to upregulate CAT protein levels (but not RNA) upon AdoMetDC inhibition, supporting the hypothesis that prozyme expression is regulated translationally. To gain insight into trans-acting factors, genetic rescue of AdoMetDC RNAi knock-down lines with human AdoMetDC was performed leading to rescue of the cell growth block, and restoration of prozyme protein to wild-type levels. Metabolite analysis showed that prozyme protein levels were inversely proportional to intracellular levels of decarboxylated AdoMet (dcAdoMet). These data suggest that prozyme translation may be regulated by dcAdoMet, a metabolite not previously identified to play a regulatory role.
© 2013 John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Metabolite-dependent regulation of gene expression in Trypanosoma brucei.Mol Microbiol. 2013 Jun;88(5):841-5. doi: 10.1111/mmi.12243. Epub 2013 May 13. Mol Microbiol. 2013. PMID: 23668674
Similar articles
-
A dual regulatory circuit consisting of S-adenosylmethionine decarboxylase protein and its reaction product controls expression of the paralogous activator prozyme in Trypanosoma brucei.PLoS Pathog. 2018 Oct 26;14(10):e1007404. doi: 10.1371/journal.ppat.1007404. eCollection 2018 Oct. PLoS Pathog. 2018. PMID: 30365568 Free PMC article.
-
Metabolite-dependent regulation of gene expression in Trypanosoma brucei.Mol Microbiol. 2013 Jun;88(5):841-5. doi: 10.1111/mmi.12243. Epub 2013 May 13. Mol Microbiol. 2013. PMID: 23668674
-
Regulated expression of an essential allosteric activator of polyamine biosynthesis in African trypanosomes.PLoS Pathog. 2008 Oct;4(10):e1000183. doi: 10.1371/journal.ppat.1000183. Epub 2008 Oct 24. PLoS Pathog. 2008. PMID: 18949025 Free PMC article.
-
Regulation and function of polyamines in African trypanosomes.Trends Parasitol. 2012 Feb;28(2):66-72. doi: 10.1016/j.pt.2011.11.001. Epub 2011 Dec 20. Trends Parasitol. 2012. PMID: 22192816 Review.
-
Translational regulation of ornithine decarboxylase and other enzymes of the polyamine pathway.Int J Biochem Cell Biol. 1999 Jan;31(1):107-22. doi: 10.1016/s1357-2725(98)00135-6. Int J Biochem Cell Biol. 1999. PMID: 10216947 Review.
Cited by
-
A dual regulatory circuit consisting of S-adenosylmethionine decarboxylase protein and its reaction product controls expression of the paralogous activator prozyme in Trypanosoma brucei.PLoS Pathog. 2018 Oct 26;14(10):e1007404. doi: 10.1371/journal.ppat.1007404. eCollection 2018 Oct. PLoS Pathog. 2018. PMID: 30365568 Free PMC article.
-
The Major Protein Arginine Methyltransferase in Trypanosoma brucei Functions as an Enzyme-Prozyme Complex.J Biol Chem. 2017 Feb 10;292(6):2089-2100. doi: 10.1074/jbc.M116.757112. Epub 2016 Dec 20. J Biol Chem. 2017. PMID: 27998975 Free PMC article.
-
Species-Selective Pyrimidineamine Inhibitors of Trypanosoma brucei S-Adenosylmethionine Decarboxylase.J Med Chem. 2018 Feb 8;61(3):1182-1203. doi: 10.1021/acs.jmedchem.7b01654. Epub 2018 Jan 5. J Med Chem. 2018. PMID: 29271204 Free PMC article.
-
Polyamine Metabolism in Leishmania Parasites: A Promising Therapeutic Target.Med Sci (Basel). 2022 Apr 22;10(2):24. doi: 10.3390/medsci10020024. Med Sci (Basel). 2022. PMID: 35645240 Free PMC article. Review.
-
Genetic validation of Trypanosoma brucei glutathione synthetase as an essential enzyme.Eukaryot Cell. 2014 May;13(5):614-24. doi: 10.1128/EC.00015-14. Epub 2014 Mar 7. Eukaryot Cell. 2014. PMID: 24610661 Free PMC article.
References
-
- Arakaki TL, Buckner FS, Gillespie JR, Malmquist NA, Phillips MA, Kalyuzhniy O, Luft JR, Detitta GT, Verlinde CL, Van Voorhis WC, Hol WG, Merritt EA. Characterization of Trypanosoma brucei dihydroorotate dehydrogenase as a possible drug target; structural, kinetic and RNAi studies. Mol Microbiol. 2008;68:37–50. - PMC - PubMed
-
- Bacchi CJ, Barker RH, Jr, Rodriguez A, Hirth B, Rattendi D, Yarlett N, Hendrick CL, Sybertz E. Trypanocidal activity of 8-methyl-5'-{[(Z)-4-aminobut-2-enyl]-(methylamino)}adenosine (Genz-644131), an adenosylmethionine decarboxylase inhibitor. Antimicrob Agents Chemother. 2009;53:3269–3272. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
