

Primary hippocampal neurons, which lack four crucial extracellular matrix molecules, display abnormalities of synaptic structure and function and severe deficits in perineuronal net formation
- PMID: 23637166
- PMCID: PMC6618965
- DOI: 10.1523/JNEUROSCI.3275-12.2013
Primary hippocampal neurons, which lack four crucial extracellular matrix molecules, display abnormalities of synaptic structure and function and severe deficits in perineuronal net formation
Abstract
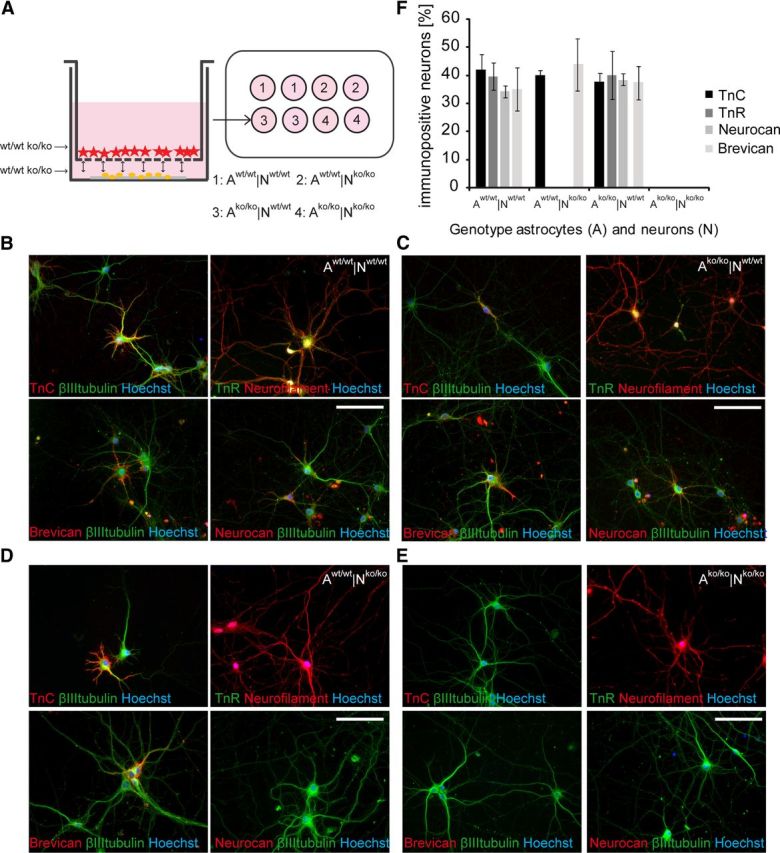
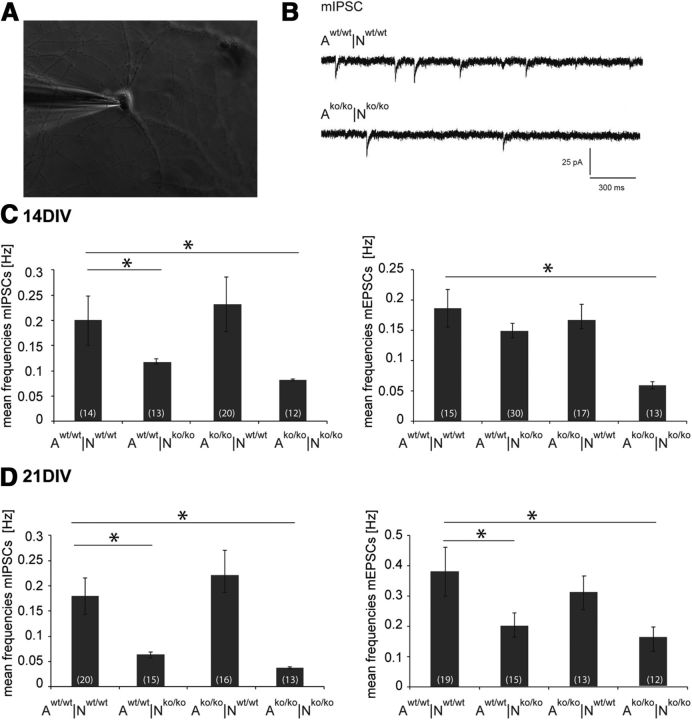
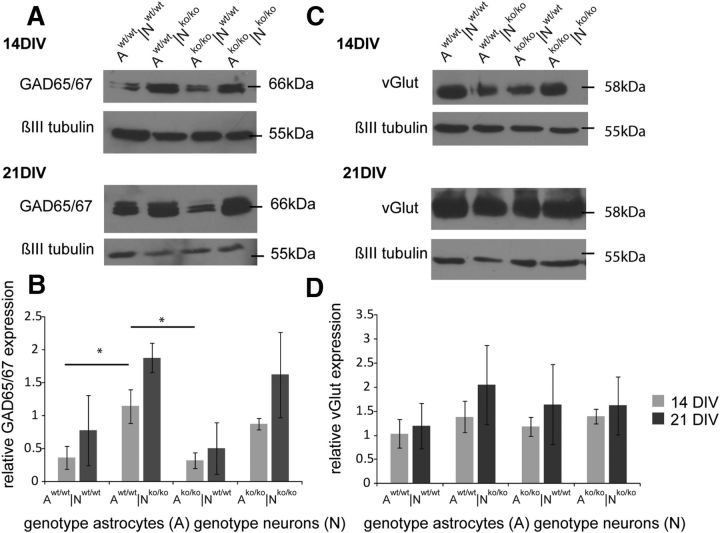
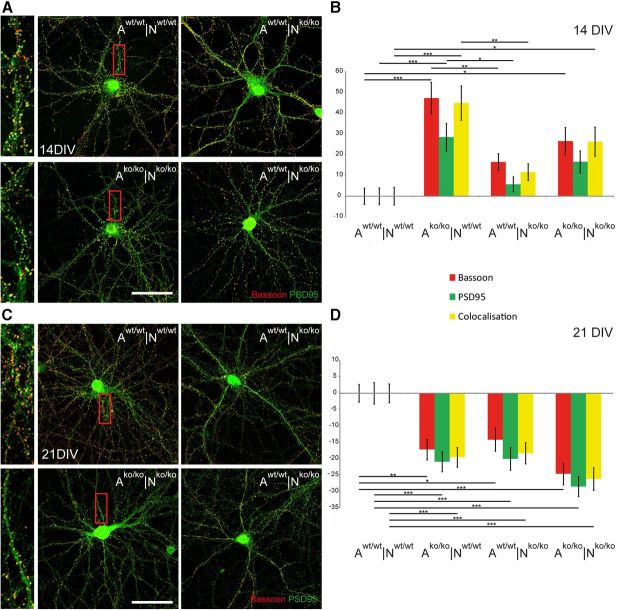
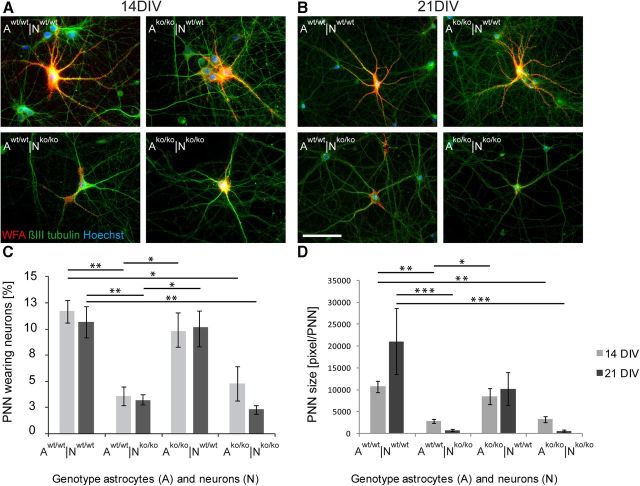
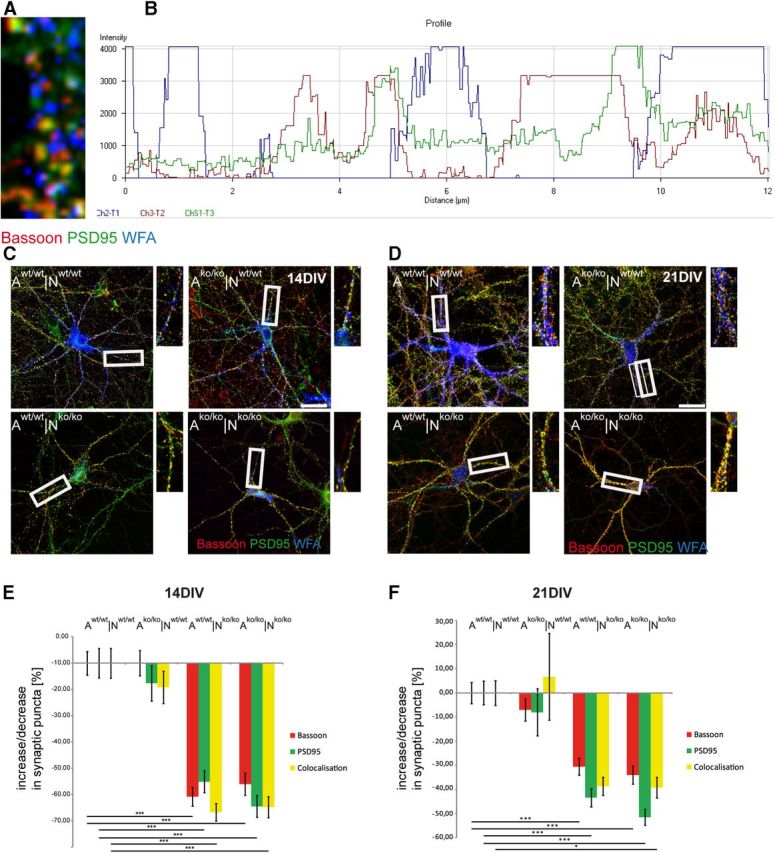
The extracellular matrix (ECM) of the brain plays crucial roles during the development, maturation, and regeneration of the CNS. In a subpopulation of neurons, the ECM condenses to superstructures called perineuronal nets (PNNs) that surround synapses. Camillo Golgi described PNNs a century ago, yet their biological functions remain elusive. Here, we studied a mouse mutant that lacks four ECM components highly enriched in the developing brain: the glycoproteins tenascin-C and tenascin-R and the chondroitin sulfate proteoglycans brevican and neurocan. Primary embryonic hippocampal neurons and astrocytes were cultivated using a cell insert system that allows for co-culture of distinct cell populations in the absence of direct membrane contacts. The wild-type and knock-out cells were combined in the four possible permutations. Using this approach, neurons cultivated in the presence of mutant astrocytes displayed a transient increase of synapses after 2 weeks. However, after a period of 3 weeks or longer, synapse formation and stabilization were compromised when either neuron or astrocyte cell populations or both were of mutant origin. The development of PNN structures was observed, but their size was substantially reduced on knock-out neurons. The synaptic activity of both wild-type and knock-out neurons was monitored using whole-cell patch clamping. The salient observation was a reduced frequency of IPSCs and EPSCs, whereas the amplitudes were not modified. Remarkably, the knock-out neuron phenotypes could not be rescued by wild-type astrocytes. We conclude that the elimination of four ECM genes compromises neuronal function.
Figures
References
-
- Brakebusch C, Seidenbecher CI, Asztely F, Rauch U, Matthies H, Meyer H, Krug M, Böckers TM, Zhou X, Kreutz MR, Montag D, Gundelfinger ED, Fässler R. Brevican-deficient mice display impaired hippocampal CA1 long-term potentiation but show no obvious deficits in learning and memory. Mol Cell Biol. 2002;22:7417–7427. doi: 10.1128/MCB.22.21.7417-7427.2002. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases