

A role for an Hsp70 nucleotide exchange factor in the regulation of synaptic vesicle endocytosis
- PMID: 23637191
- PMCID: PMC3707978
- DOI: 10.1523/JNEUROSCI.4505-12.2013
A role for an Hsp70 nucleotide exchange factor in the regulation of synaptic vesicle endocytosis
Abstract
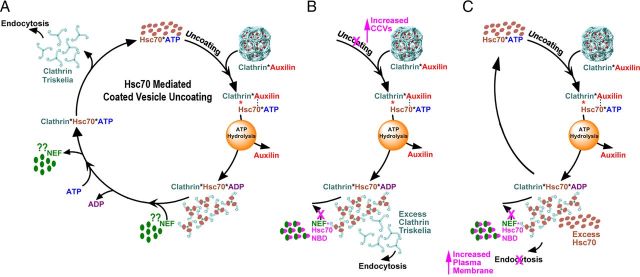
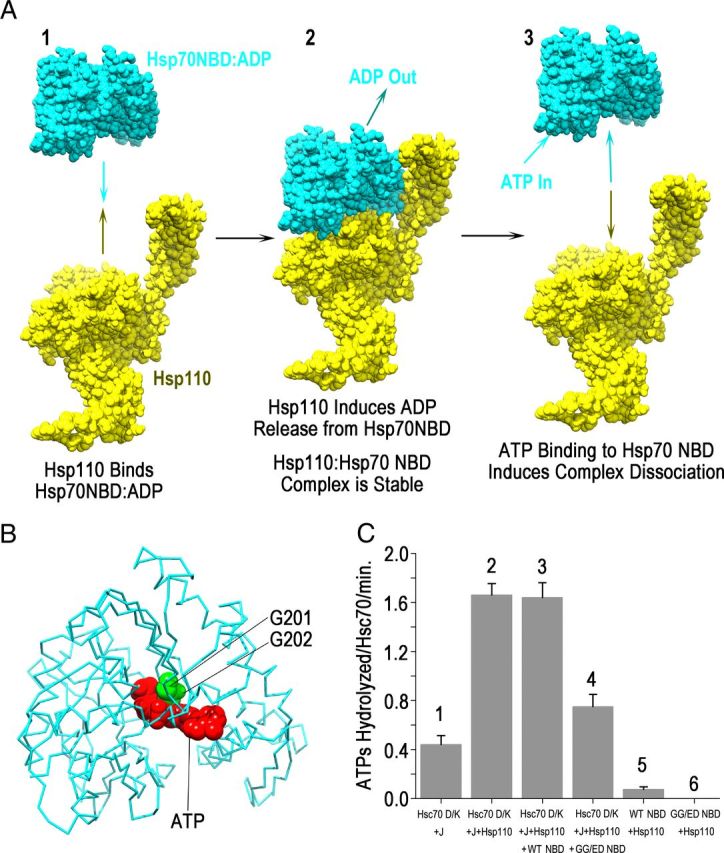
Neurotransmission requires a continuously available pool of synaptic vesicles (SVs) that can fuse with the plasma membrane and release their neurotransmitter contents upon stimulation. After fusion, SV membranes and membrane proteins are retrieved from the presynaptic plasma membrane by clathrin-mediated endocytosis. After the internalization of a clathrin-coated vesicle, the vesicle must uncoat to replenish the pool of SVs. Clathrin-coated vesicle uncoating requires ATP and is mediated by the ubiquitous molecular chaperone Hsc70. In vitro, depolymerized clathrin forms a stable complex with Hsc70*ADP. This complex can be dissociated by nucleotide exchange factors (NEFs) that release ADP from Hsc70, allowing ATP to bind and induce disruption of the clathrin:Hsc70 association. Whether NEFs generally play similar roles in vesicle trafficking in vivo and whether they play such roles in SV endocytosis in particular is unknown. To address this question, we used information from recent structural and mechanistic studies of Hsp70:NEF and Hsp70:co-chaperone interactions to design a NEF inhibitor. Using acute perturbations at giant reticulospinal synapses of the sea lamprey (Petromyzon marinus), we found that this NEF inhibitor inhibited SV endocytosis. When this inhibitor was mutated so that it could no longer bind and inhibit Hsp110 (a NEF that we find to be highly abundant in brain cytosol), its ability to inhibit SV endocytosis was eliminated. These observations indicate that the action of a NEF, most likely Hsp110, is normally required during SV trafficking to release clathrin from Hsc70 and make it available for additional rounds of endocytosis.
Figures






Similar articles
-
The role of molecular chaperones in clathrin mediated vesicular trafficking.Front Mol Biosci. 2015 May 19;2:26. doi: 10.3389/fmolb.2015.00026. eCollection 2015. Front Mol Biosci. 2015. PMID: 26042225 Free PMC article. Review.
-
Hsc70 chaperones clathrin and primes it to interact with vesicle membranes.J Biol Chem. 2000 Mar 24;275(12):8439-47. doi: 10.1074/jbc.275.12.8439. J Biol Chem. 2000. PMID: 10722678
-
Clathrin and synaptic vesicle endocytosis: studies at the squid giant synapse.Biochem Soc Trans. 2006 Feb;34(Pt 1):68-72. doi: 10.1042/BST0340068. Biochem Soc Trans. 2006. PMID: 16417485 Free PMC article. Review.
-
Dominant-interfering Hsc70 mutants disrupt multiple stages of the clathrin-coated vesicle cycle in vivo.J Cell Biol. 2001 Feb 5;152(3):607-20. doi: 10.1083/jcb.152.3.607. J Cell Biol. 2001. PMID: 11157986 Free PMC article.
-
Hsp110 is a nucleotide-activated exchange factor for Hsp70.J Biol Chem. 2008 Apr 4;283(14):8877-84. doi: 10.1074/jbc.M710063200. Epub 2008 Jan 24. J Biol Chem. 2008. PMID: 18218635
Cited by
-
Differential regulation of synaptic AP-2/clathrin vesicle uncoating in synaptic plasticity.Sci Rep. 2017 Nov 17;7(1):15781. doi: 10.1038/s41598-017-16055-4. Sci Rep. 2017. PMID: 29150658 Free PMC article.
-
Structural mechanisms of chaperone mediated protein disaggregation.Front Mol Biosci. 2014 Sep 15;1:12. doi: 10.3389/fmolb.2014.00012. eCollection 2014. Front Mol Biosci. 2014. PMID: 25988153 Free PMC article. Review.
-
Clathrin coat controls synaptic vesicle acidification by blocking vacuolar ATPase activity.Elife. 2018 Apr 13;7:e32569. doi: 10.7554/eLife.32569. Elife. 2018. PMID: 29652249 Free PMC article.
-
J protein mutations and resulting proteostasis collapse.Front Cell Neurosci. 2014 Jul 8;8:191. doi: 10.3389/fncel.2014.00191. eCollection 2014. Front Cell Neurosci. 2014. PMID: 25071450 Free PMC article. Review.
-
The role of molecular chaperones in clathrin mediated vesicular trafficking.Front Mol Biosci. 2015 May 19;2:26. doi: 10.3389/fmolb.2015.00026. eCollection 2015. Front Mol Biosci. 2015. PMID: 26042225 Free PMC article. Review.
References
-
- Blondeau F, Ritter B, Allaire PD, Wasiak S, Girard M, Hussain NK, Angers A, Legendre-Guillemin V, Roy L, Boismenu D, Kearney RE, Bell AW, Bergeron JJ, McPherson PS. Tandem MS analysis of brain clathrin-coated vesicles reveals their critical involvement in synaptic vesicle recycling. Proc Natl Acad Sci U S A. 2004;101:3833–3838. doi: 10.1073/pnas.0308186101. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous