

cAMP-mediated stabilization of fusion pores in cultured rat pituitary lactotrophs
- PMID: 23637196
- PMCID: PMC3674111
- DOI: 10.1523/JNEUROSCI.5351-12.2013
cAMP-mediated stabilization of fusion pores in cultured rat pituitary lactotrophs
Abstract
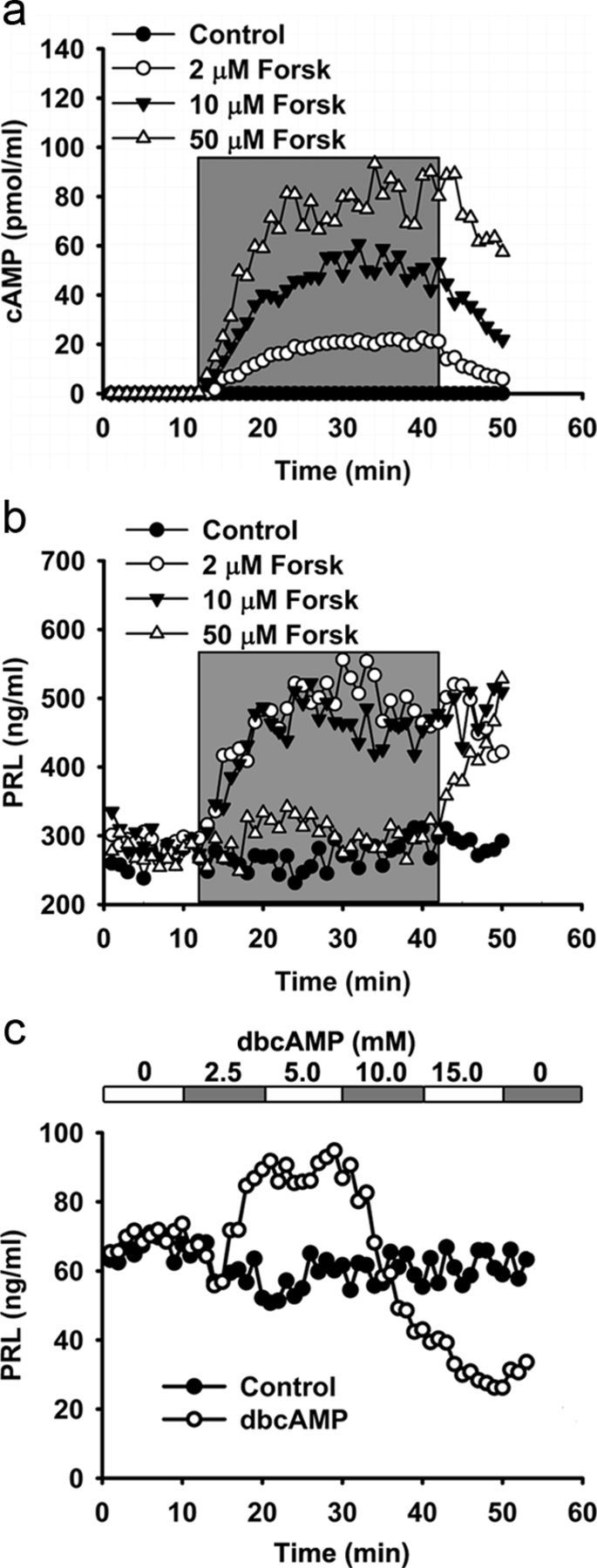
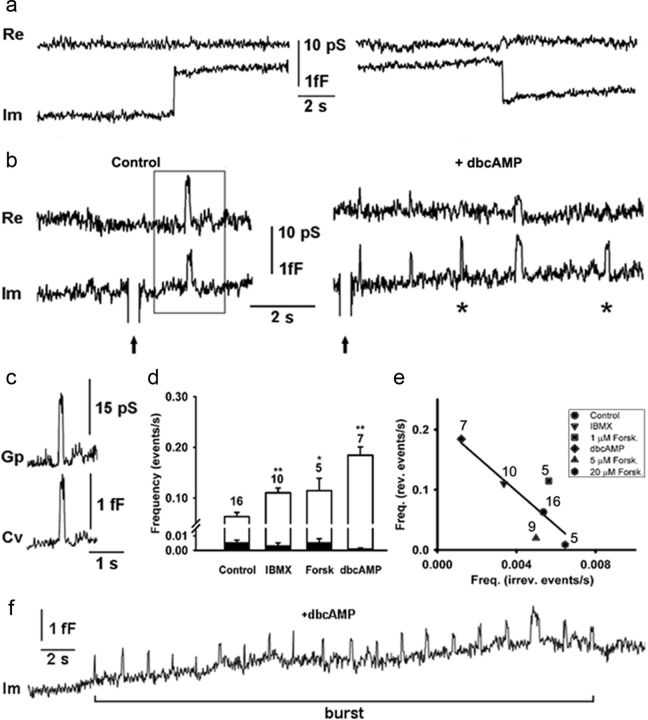
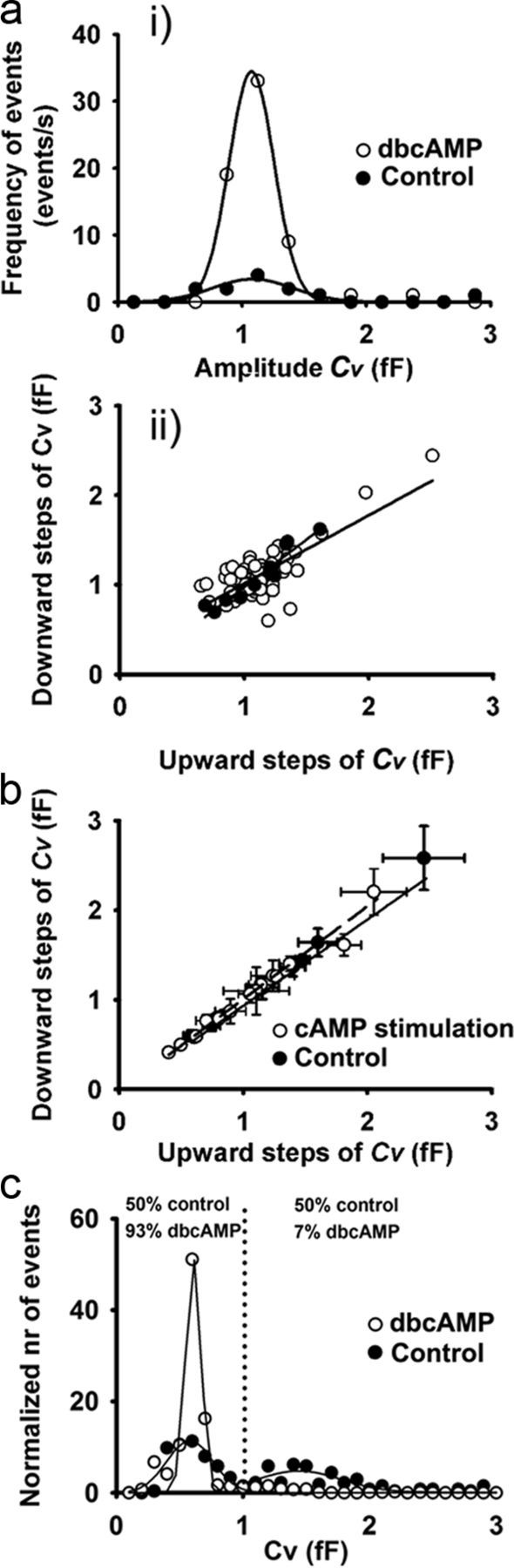
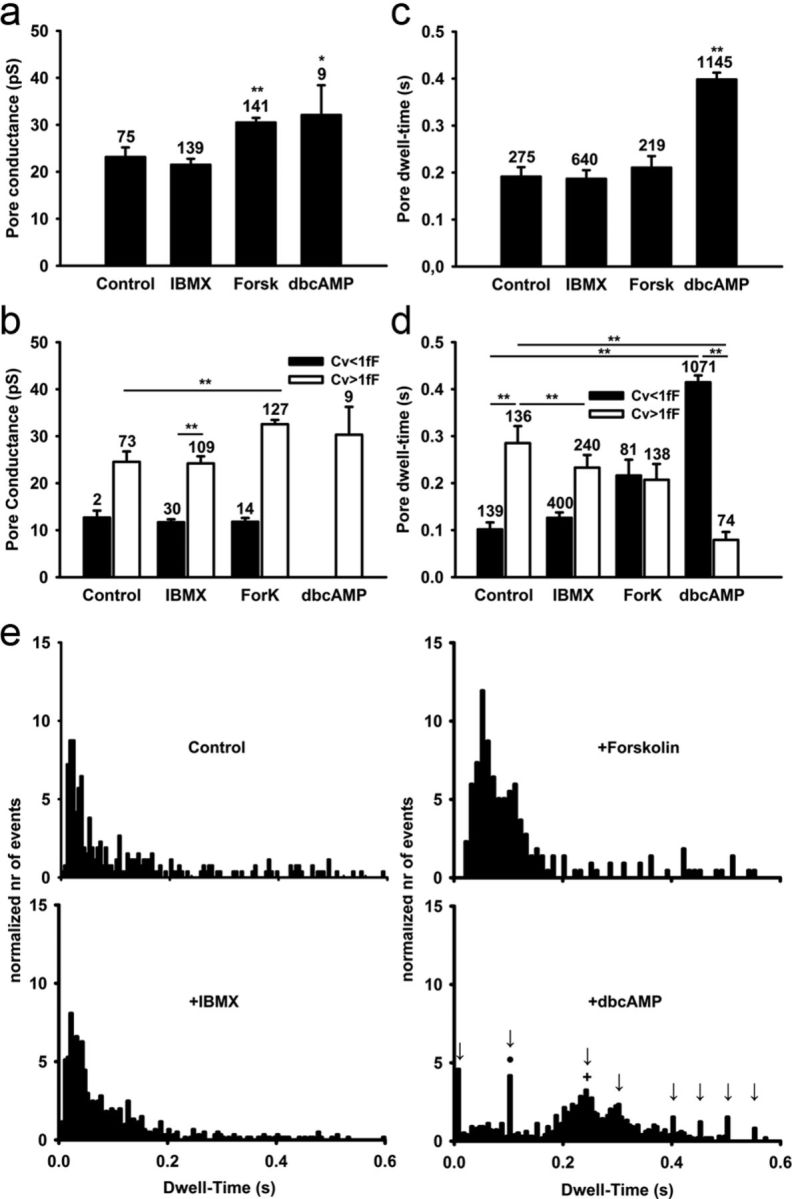
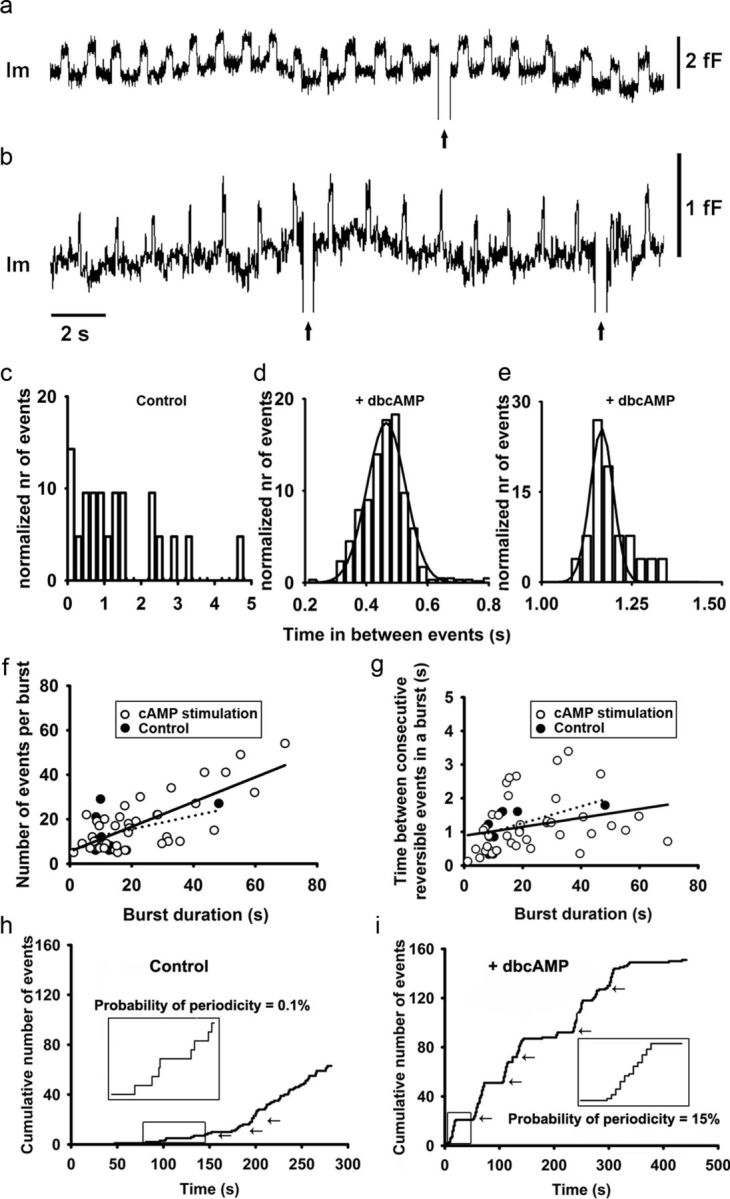
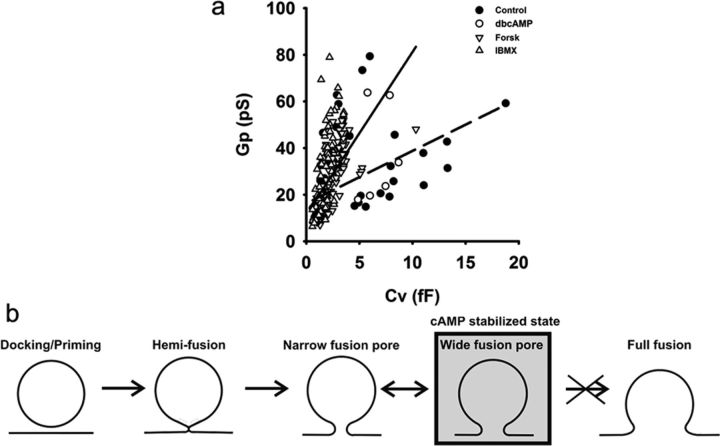
Regulated exocytosis mediates the release of hormones and transmitters. The last step of this process is represented by the merger between the vesicle and the plasma membranes, and the formation of a fusion pore. Once formed, the initially stable and narrow fusion pore may reversibly widen (transient exocytosis) or fully open (full-fusion exocytosis). Exocytosis is typically triggered by an elevation in cytosolic calcium activity. However, other second messengers, such as cAMP, have been reported to modulate secretion. The way in which cAMP influences the transitions between different fusion pore states remains unclear. Here, hormone release studies show that prolactin release from isolated rat lactotrophs stimulated by forskolin, an activator of adenylyl cyclases, and by membrane-permeable cAMP analog (dbcAMP), exhibit a biphasic concentration dependency. Although at lower concentrations (2-10 μm forskolin and 2.5-5 mm dbcAMP) these agents stimulate prolactin release, an inhibition is measured at higher concentrations (50 μm forskolin and 10-15 mm dbcAMP). By using high-resolution capacitance (Cm) measurements, we recorded discrete increases in Cm, which represent elementary exocytic events. An elevation of cAMP leaves the frequency of full-fusion events unchanged while increasing the frequency of transient events. These exhibited a wider fusion pore as measured by increased fusion pore conductance and a prolonged fusion pore dwell time. The probability of observing rhythmic reopening of transient fusion pores was elevated by dbcAMP. In conclusion, cAMP-mediated stabilization of wide fusion pores prevents vesicles from proceeding to the full-fusion stage of exocytosis, which hinders vesicle content discharge at high cAMP concentrations.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources