

Phosphorylation of centromeric histone H3 variant regulates chromosome segregation in Saccharomyces cerevisiae
- PMID: 23637466
- PMCID: PMC3681705
- DOI: 10.1091/mbc.E12-12-0893
Phosphorylation of centromeric histone H3 variant regulates chromosome segregation in Saccharomyces cerevisiae
Abstract
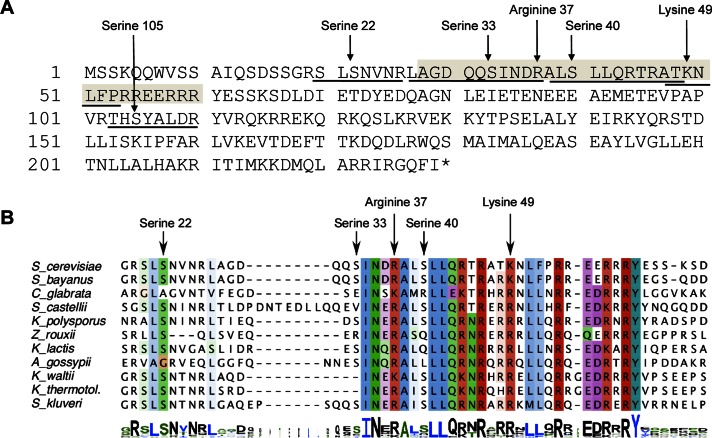
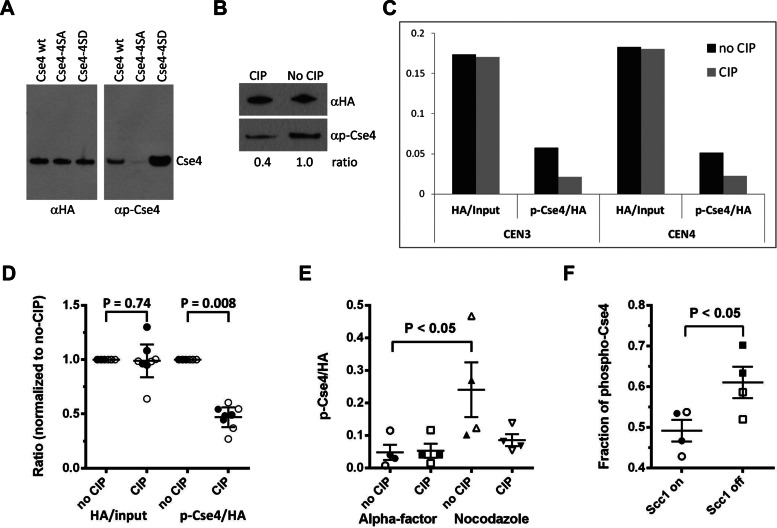
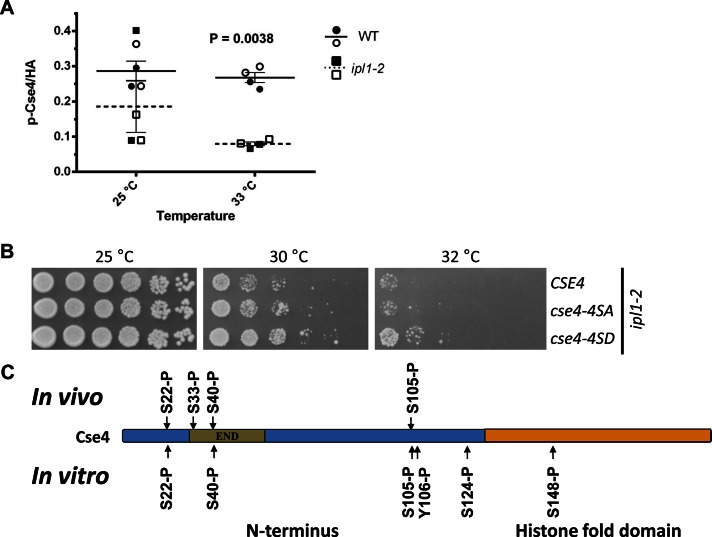
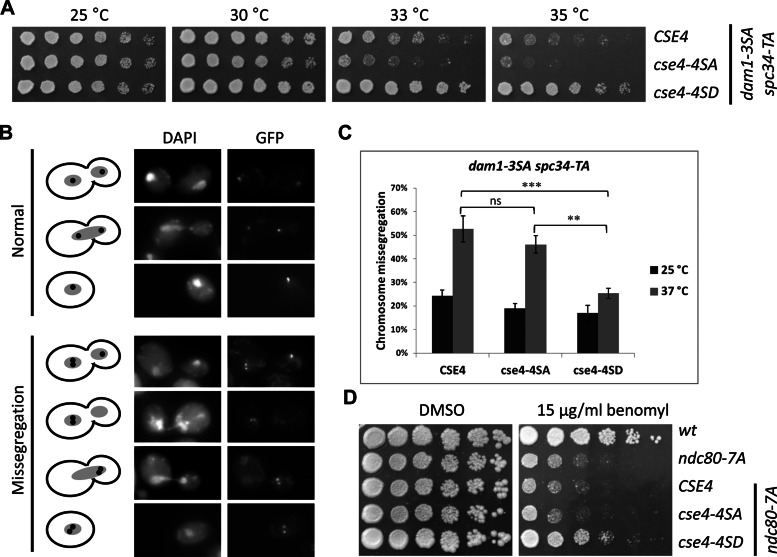
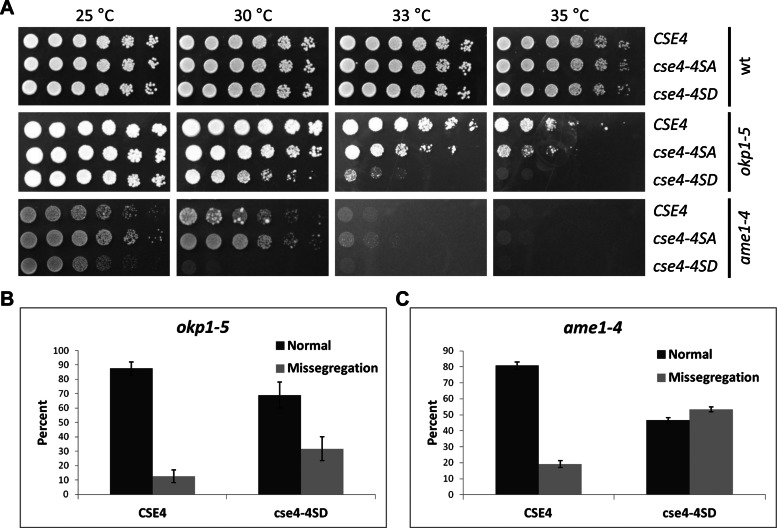
The centromeric histone H3 variant (CenH3) is essential for chromosome segregation in eukaryotes. We identify posttranslational modifications of Saccharomyces cerevisiae CenH3, Cse4. Functional characterization of cse4 phosphorylation mutants shows growth and chromosome segregation defects when combined with kinetochore mutants okp1 and ame1. Using a phosphoserine-specific antibody, we show that the association of phosphorylated Cse4 with centromeres increases in response to defective microtubule attachment or reduced cohesion. We determine that evolutionarily conserved Ipl1/Aurora B contributes to phosphorylation of Cse4, as levels of phosphorylated Cse4 are reduced at centromeres in ipl1 strains in vivo, and in vitro assays show phosphorylation of Cse4 by Ipl1. Consistent with these results, we observe that a phosphomimetic cse4-4SD mutant suppresses the temperature-sensitive growth of ipl1-2 and Ipl1 substrate mutants dam1 spc34 and ndc80, which are defective for chromosome biorientation. Furthermore, cell biology approaches using a green fluorescent protein-labeled chromosome show that cse4-4SD suppresses chromosome segregation defects in dam1 spc34 strains. On the basis of these results, we propose that phosphorylation of Cse4 destabilizes defective kinetochores to promote biorientation and ensure faithful chromosome segregation. Taken together, our results provide a detailed analysis, in vivo and in vitro, of Cse4 phosphorylation and its role in promoting faithful chromosome segregation.
Figures
References
-
- Black BE, Jansen LE, Maddox PS, Foltz DR, Desai AB, Shah JV, Cleveland DW. Centromere identity maintained by nucleosomes assembled with histone H3 containing the CENP-A targeting domain. Mol Cell. 2007;25:309–322. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
