

feedback between population and evolutionary dynamics determines the fate of social microbial populations
- PMID: 23637571
- PMCID: PMC3640081
- DOI: 10.1371/journal.pbio.1001547
feedback between population and evolutionary dynamics determines the fate of social microbial populations
Abstract
The evolutionary spread of cheater strategies can destabilize populations engaging in social cooperative behaviors, thus demonstrating that evolutionary changes can have profound implications for population dynamics. At the same time, the relative fitness of cooperative traits often depends upon population density, thus leading to the potential for bi-directional coupling between population density and the evolution of a cooperative trait. Despite the potential importance of these eco-evolutionary feedback loops in social species, they have not yet been demonstrated experimentally and their ecological implications are poorly understood. Here, we demonstrate the presence of a strong feedback loop between population dynamics and the evolutionary dynamics of a social microbial gene, SUC2, in laboratory yeast populations whose cooperative growth is mediated by the SUC2 gene. We directly visualize eco-evolutionary trajectories of hundreds of populations over 50-100 generations, allowing us to characterize the phase space describing the interplay of evolution and ecology in this system. Small populations collapse despite continual evolution towards increased cooperative allele frequencies; large populations with a sufficient number of cooperators "spiral" to a stable state of coexistence between cooperator and cheater strategies. The presence of cheaters does not significantly affect the equilibrium population density, but it does reduce the resilience of the population as well as its ability to adapt to a rapidly deteriorating environment. Our results demonstrate the potential ecological importance of coupling between evolutionary dynamics and the population dynamics of cooperatively growing organisms, particularly in microbes. Our study suggests that this interaction may need to be considered in order to explain intraspecific variability in cooperative behaviors, and also that this feedback between evolution and ecology can critically affect the demographic fate of those species that rely on cooperation for their survival.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
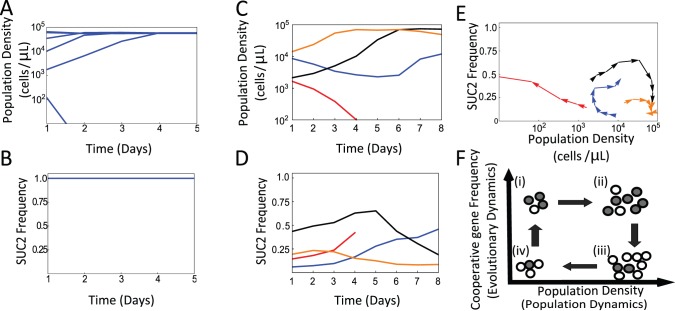

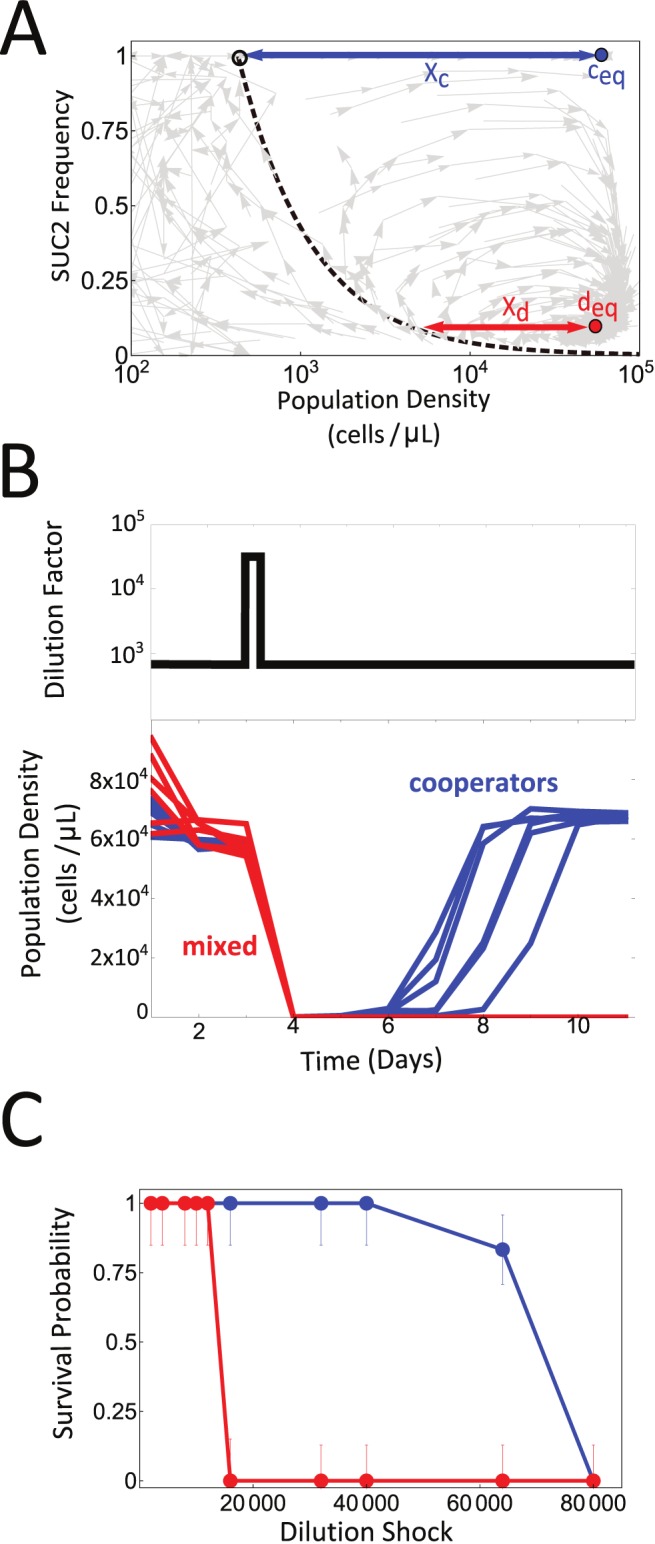
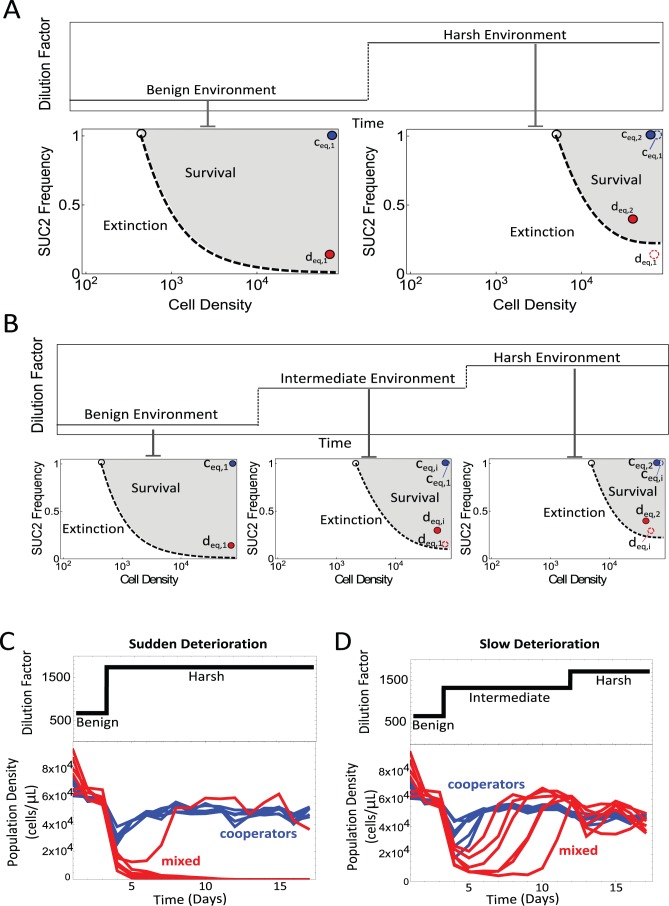
Comment in
-
Cooperation and the fate of microbial societies.PLoS Biol. 2013;11(4):e1001549. doi: 10.1371/journal.pbio.1001549. Epub 2013 Apr 30. PLoS Biol. 2013. PMID: 23637573 Free PMC article.
References
-
- Schoener TW (2011) The newest synthesis: understanding the interplay of evolutionary and ecological dynamics. Science 331: 426–429. - PubMed
-
- Hastings A (1997) Population biology. New York: Springer.
-
- Grant PR, Grant BR (2006) Evolution of character displacement in Darwin's finches. Science 313: 224–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
