

Sex-differential selection and the evolution of X inactivation strategies
- PMID: 23637618
- PMCID: PMC3630082
- DOI: 10.1371/journal.pgen.1003440
Sex-differential selection and the evolution of X inactivation strategies
Abstract
X inactivation--the transcriptional silencing of one X chromosome copy per female somatic cell--is universal among therian mammals, yet the choice of which X to silence exhibits considerable variation among species. X inactivation strategies can range from strict paternally inherited X inactivation (PXI), which renders females haploid for all maternally inherited alleles, to unbiased random X inactivation (RXI), which equalizes expression of maternally and paternally inherited alleles in each female tissue. However, the underlying evolutionary processes that might account for this observed diversity of X inactivation strategies remain unclear. We present a theoretical population genetic analysis of X inactivation evolution and specifically consider how conditions of dominance, linkage, recombination, and sex-differential selection each influence evolutionary trajectories of X inactivation. The results indicate that a single, critical interaction between allelic dominance and sex-differential selection can select for a broad and continuous range of X inactivation strategies, including unequal rates of inactivation between maternally and paternally inherited X chromosomes. RXI is favored over complete PXI as long as alleles deleterious to female fitness are sufficiently recessive, and the criteria for RXI evolution is considerably more restrictive when fitness variation is sexually antagonistic (i.e., alleles deleterious to females are beneficial to males) relative to variation that is deleterious to both sexes. Evolutionary transitions from PXI to RXI also generally increase mean relative female fitness at the expense of decreased male fitness. These results provide a theoretical framework for predicting and interpreting the evolution of chromosome-wide expression of X-linked genes and lead to several useful predictions that could motivate future studies of allele-specific gene expression variation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
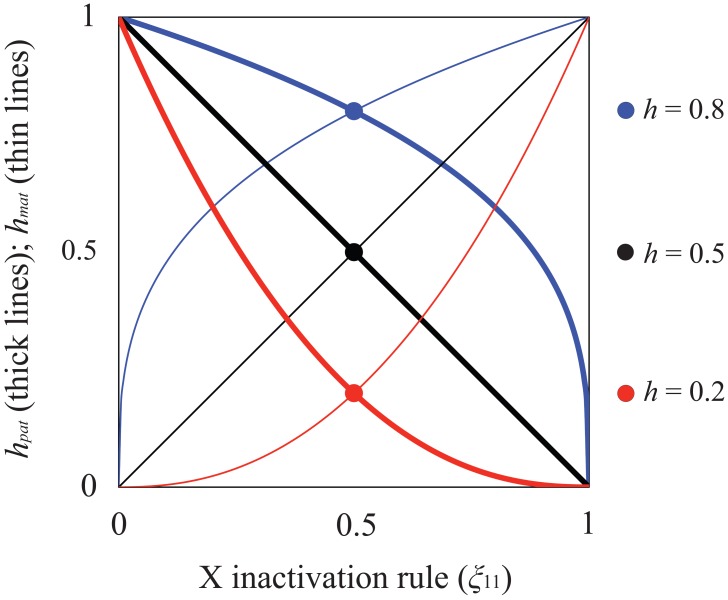
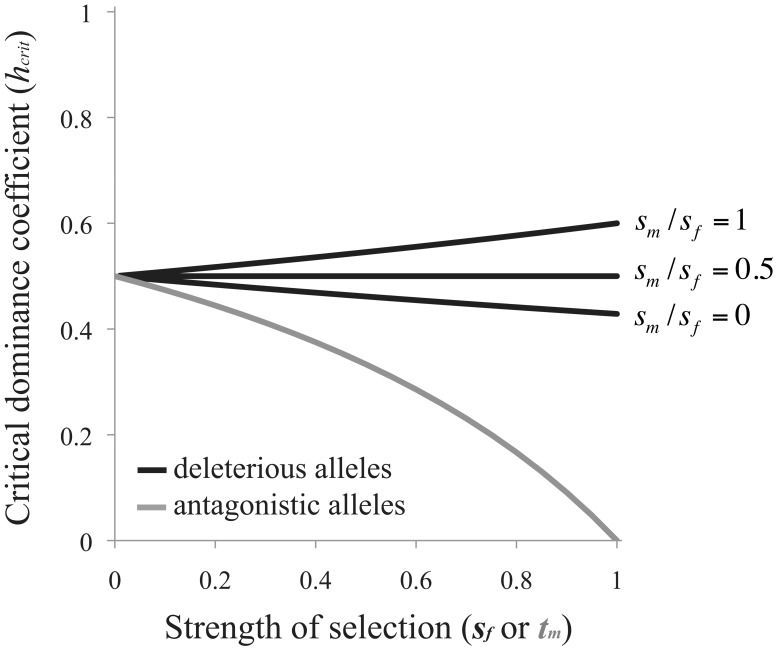
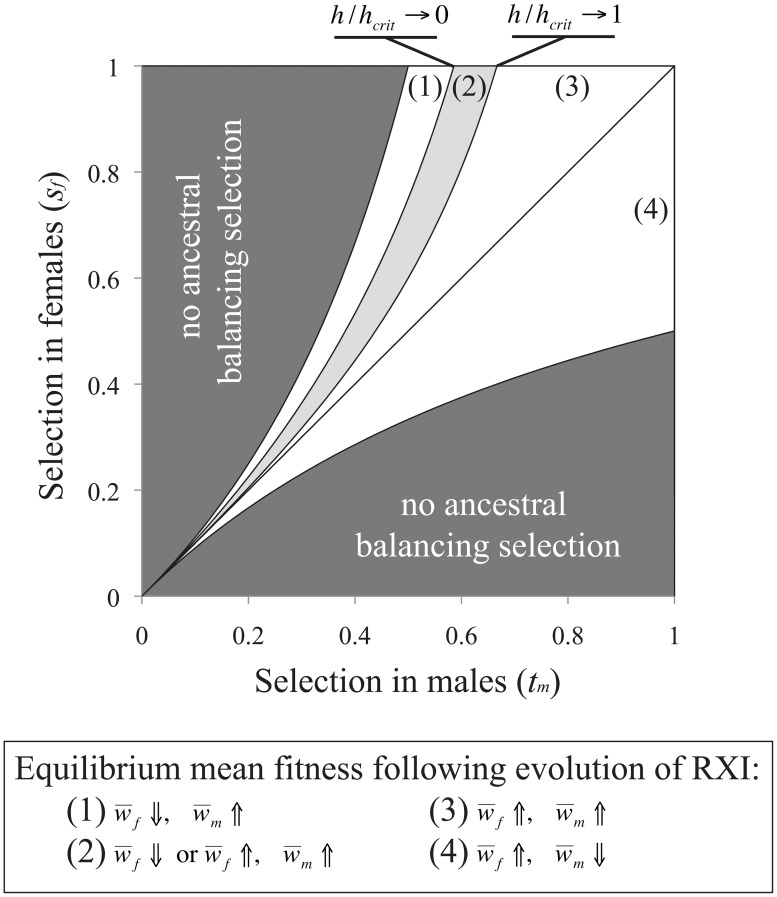
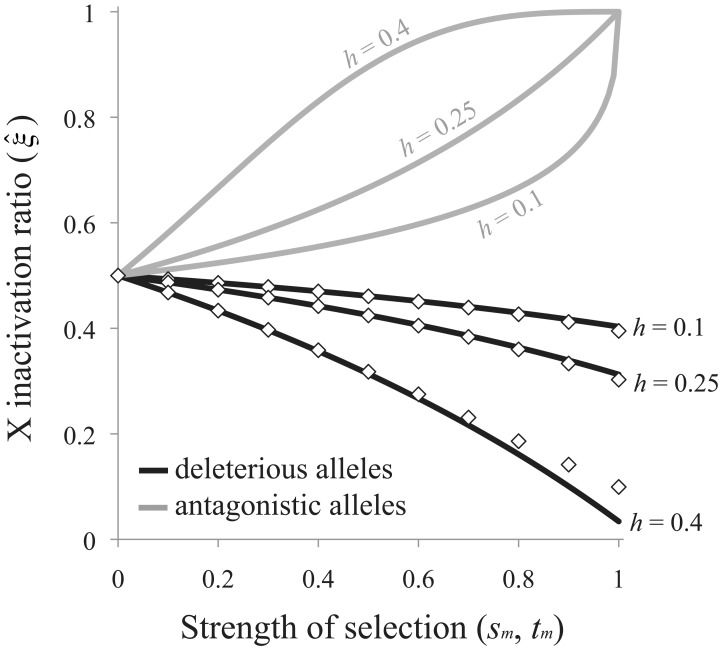
Similar articles
-
Self-imposed silence: parental antagonism and the evolution of X-chromosome inactivation.Evolution. 2006 Mar;60(3):440-7. Evolution. 2006. PMID: 16637489
-
Genomic imprinting leads to less selectively maintained polymorphism on X chromosomes.Genetics. 2012 Dec;192(4):1455-64. doi: 10.1534/genetics.112.145607. Epub 2012 Sep 28. Genetics. 2012. PMID: 23023005 Free PMC article.
-
Sexual antagonism and the evolution of X chromosome inactivation.Evolution. 2008 Aug;62(8):2097-104. doi: 10.1111/j.1558-5646.2008.00431.x. Epub 2008 May 27. Evolution. 2008. PMID: 18507742
-
Origin and evolution of X chromosome inactivation.Curr Opin Cell Biol. 2012 Jun;24(3):397-404. doi: 10.1016/j.ceb.2012.02.004. Epub 2012 Mar 14. Curr Opin Cell Biol. 2012. PMID: 22425180 Review.
-
Different flavors of X-chromosome inactivation in mammals.Curr Opin Cell Biol. 2013 Jun;25(3):314-21. doi: 10.1016/j.ceb.2013.03.001. Epub 2013 Apr 8. Curr Opin Cell Biol. 2013. PMID: 23578369 Review.
Cited by
-
Evidence for increased levels of positive and negative selection on the X chromosome versus autosomes in humans.Mol Biol Evol. 2014 Sep;31(9):2267-82. doi: 10.1093/molbev/msu166. Epub 2014 May 15. Mol Biol Evol. 2014. PMID: 24830675 Free PMC article.
-
Retrogene Duplication and Expression Patterns Shaped by the Evolution of Sex Chromosomes in Malaria Mosquitoes.Genes (Basel). 2022 May 28;13(6):968. doi: 10.3390/genes13060968. Genes (Basel). 2022. PMID: 35741730 Free PMC article.
-
The evolution of X chromosome inactivation in mammals: the demise of Ohno's hypothesis?Cell Mol Life Sci. 2014 Apr;71(8):1383-94. doi: 10.1007/s00018-013-1499-6. Epub 2013 Oct 31. Cell Mol Life Sci. 2014. PMID: 24173285 Free PMC article. Review.
-
Plastin 3 in health and disease: a matter of balance.Cell Mol Life Sci. 2021 Jul;78(13):5275-5301. doi: 10.1007/s00018-021-03843-5. Epub 2021 May 23. Cell Mol Life Sci. 2021. PMID: 34023917 Free PMC article. Review.
-
A novel mutation in PLS3 causes extremely rare X-linked osteogenesis imperfecta.Mol Genet Genomic Med. 2020 Dec;8(12):e1525. doi: 10.1002/mgg3.1525. Epub 2020 Nov 9. Mol Genet Genomic Med. 2020. PMID: 33166085 Free PMC article.
References
-
- Payer B, Lee JT (2008) X chromosome dosage compensation: how mammals keep the balance. Ann Rev Genet 42: 733–772. - PubMed
-
- Deakin JE, Chaumeil J, Hore TA, Graves JAM (2009) Unravelling the evolutionary origins of X chromosome inactivation in mammals: insights from marsupials and monotremes. Chromosome Research 17: 671–685. - PubMed
-
- Cooper DW, VandeBerg JL, Sharman GB, Poole WE (1971) Phosphoglycerate kinase polymorphism in kangaroos provides further evidence for paternal X inactivation. Nature New Biology 230: 155–157. - PubMed
-
- Sharman GB (1971) Late DNA replication in the paternally derived X chromosome of female kangaroos. Nature 230: 231–232. - PubMed
-
- Lyon MF (1961) Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature 190: 372–373. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
