

Genome-wide association studies identify two novel BMP15 mutations responsible for an atypical hyperprolificacy phenotype in sheep
- PMID: 23637641
- PMCID: PMC3636084
- DOI: 10.1371/journal.pgen.1003482
Genome-wide association studies identify two novel BMP15 mutations responsible for an atypical hyperprolificacy phenotype in sheep
Abstract
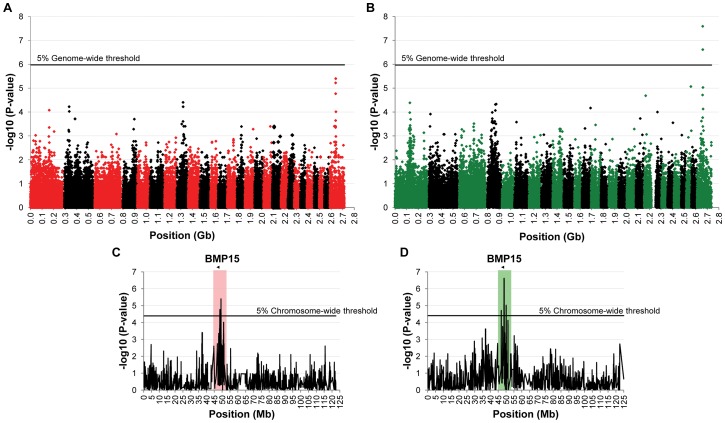
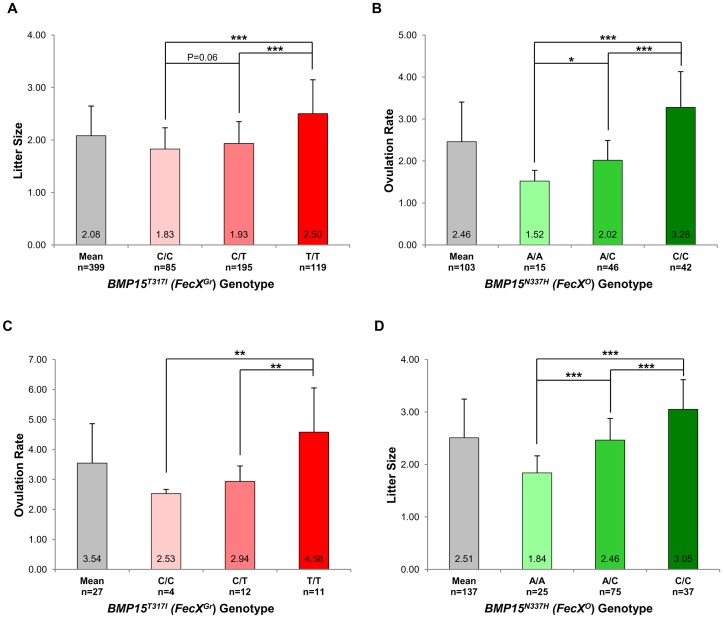
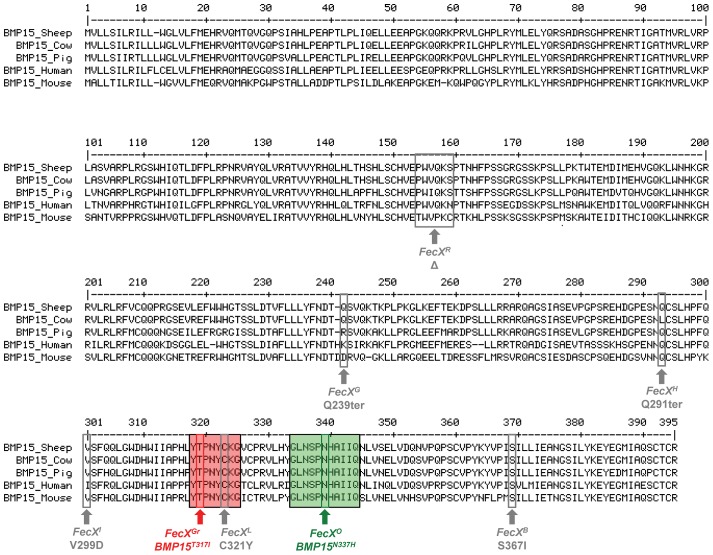
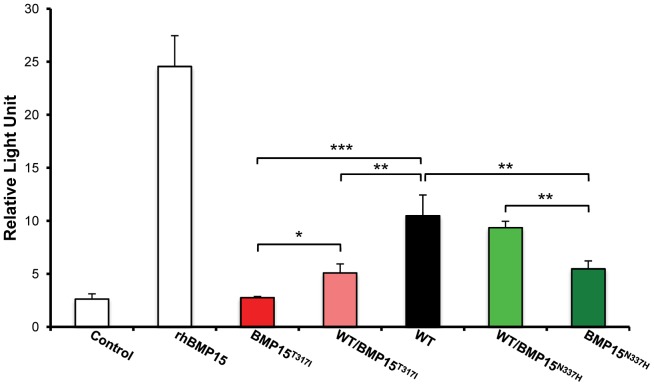
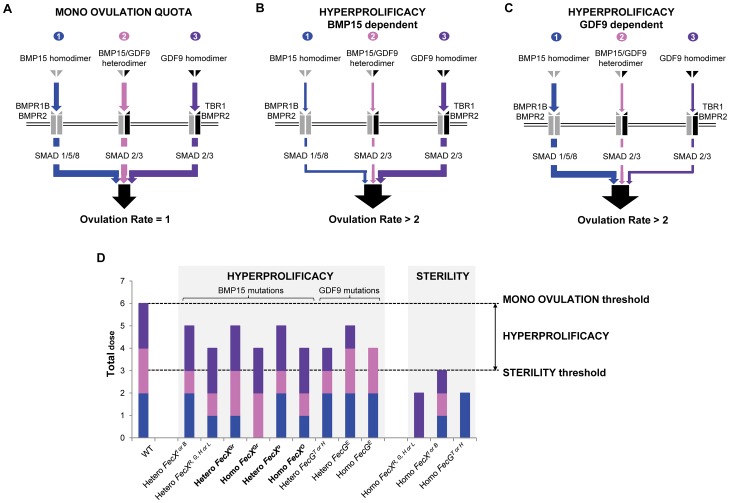
Some sheep breeds are naturally prolific, and they are very informative for the studies of reproductive genetics and physiology. Major genes increasing litter size (LS) and ovulation rate (OR) were suspected in the French Grivette and the Polish Olkuska sheep populations, respectively. To identify genetic variants responsible for the highly prolific phenotype in these two breeds, genome-wide association studies (GWAS) followed by complementary genetic and functional analyses were performed. Highly prolific ewes (cases) and normal prolific ewes (controls) from each breed were genotyped using the Illumina OvineSNP50 Genotyping Beadchip. In both populations, an X chromosome region, close to the BMP15 gene, harbored clusters of markers with suggestive evidence of association at significance levels between 1E(-05) and 1E(-07). The BMP15 candidate gene was then sequenced, and two novel non-conservative mutations called FecX(Gr) and FecX(O) were identified in the Grivette and Olkuska breeds, respectively. The two mutations were associated with the highly prolific phenotype (p FecX (Gr) = 5.98E(-06) and p FecX(O) = 2.55E(-08)). Homozygous ewes for the mutated allele showed a significantly increased prolificacy (FecX(Gr)/FecX(Gr), LS = 2.50 ± 0.65 versus FecX(+)/FecX(Gr), LS = 1.93 ± 0.42, p<1E(-03) and FecX(O)/FecX(O), OR = 3.28 ± 0.85 versus FecX(+)/FecX(O), OR = 2.02 ± 0.47, p<1E(-03)). Both mutations are located in very well conserved motifs of the protein and altered the BMP15 signaling activity in vitro using a BMP-responsive luciferase test in COV434 granulosa cells. Thus, we have identified two novel mutations in the BMP15 gene associated with increased LS and OR. Notably, homozygous FecX(Gr)/FecX(Gr) Grivette and homozygous FecX(O)/FecX(O) Olkuska ewes are hyperprolific in striking contrast with the sterility exhibited by all other known homozygous BMP15 mutations. Our results bring new insights into the key role played by the BMP15 protein in ovarian function and could contribute to a better understanding of the pathogenesis of women's fertility disorders.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Matzuk MM, Burns KH, Viveiros MM, Eppig JJ (2002) Intercellular communication in the mammalian ovary: oocytes carry the conversation. Science 296: 2178–2180. - PubMed
-
- Gilchrist RB, Lane M, Thompson JG (2008) Oocyte-secreted factors: regulators of cumulus cell function and oocyte quality. Hum Reprod Update 14: 159–177. - PubMed
-
- Dong J, Albertini DF, Nishimori K, Kumar TR, Lu N, et al. (1996) Growth differentiation factor-9 is required during early ovarian folliculogenesis. Nature 383: 531–535. - PubMed
-
- Nilsson EE, Skinner MK (2002) Growth and differentiation factor-9 stimulates progression of early primary but not primordial rat ovarian follicle development. Biol Reprod 67: 1018–1024. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
