

Astroglial networking contributes to neurometabolic coupling
- PMID: 23637659
- PMCID: PMC3636502
- DOI: 10.3389/fnene.2013.00004
Astroglial networking contributes to neurometabolic coupling
Abstract
The strategic position of astrocytic processes between blood capillaries and neurons, provided the early insight that astrocytes play a key role in supplying energy substrates to neurons in an activity-dependent manner. The central role of astrocytes in neurometabolic coupling has been first established at the level of single cell. Since then, exciting recent work based on cellular imaging and electrophysiological recordings has provided new mechanistic insights into this phenomenon, revealing the crucial role of gap junction (GJ)-mediated networks of astrocytes. Indeed, astrocytes define the local availability of energy substrates by regulating blood flow. Subsequently, in order to efficiently reach distal neurons, these substrates can be taken up, and distributed through networks of astrocytes connected by GJs, a process modulated by neuronal activity. Astrocytic networks can be morphologically and/or functionally altered in the course of various pathological conditions, raising the intriguing possibility of a direct contribution from these networks to neuronal dysfunction. The present review upgrades the current view of neuroglial metabolic coupling, by including the recently unravelled properties of astroglial metabolic networks and their potential contribution to normal and pathological neuronal activity.
Keywords: astrocytes; astroglial networks; energy metabolism; epilepsy; gap junctions; neurodegenerative diseases; neuroglial interactions; neurometabolic coupling.
Figures
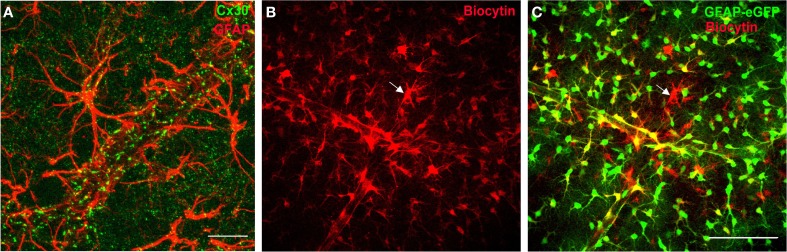
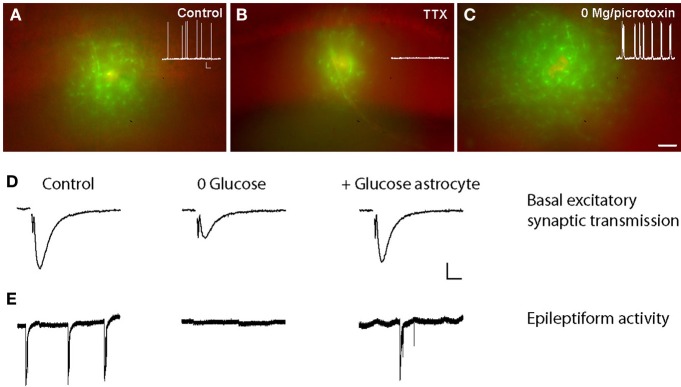
References
-
- Ball K. K., Gandhi G. K., Thrash J., Cruz N. F., Dienel G. A. (2007). Astrocytic connexin distributions and rapid, extensive dye transfer via gap junctions in the inferior colliculus: implications for [(14)C]glucose metabolite trafficking. J. Neurosci. Res. 85, 3267–3283 10.1002/jnr.21376 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous