

Absence of complementary sex determination in the parasitoid wasp genus Asobara (Hymenoptera: Braconidae)
- PMID: 23637750
- PMCID: PMC3614920
- DOI: 10.1371/journal.pone.0060459
Absence of complementary sex determination in the parasitoid wasp genus Asobara (Hymenoptera: Braconidae)
Abstract
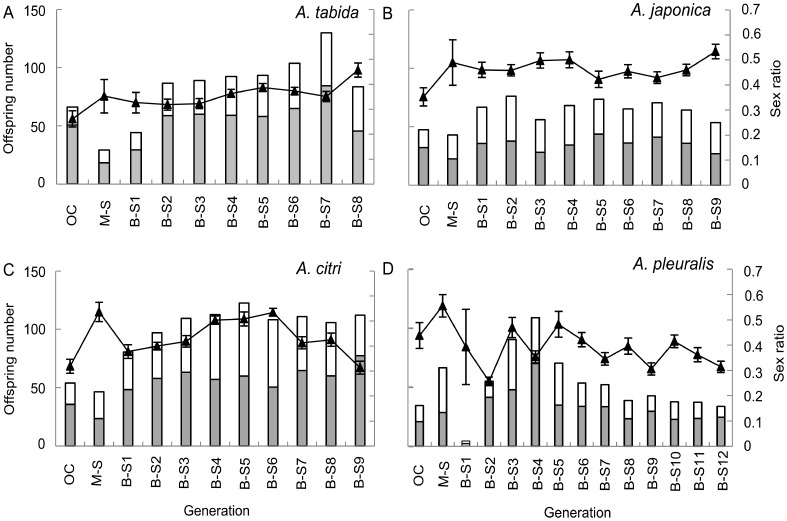
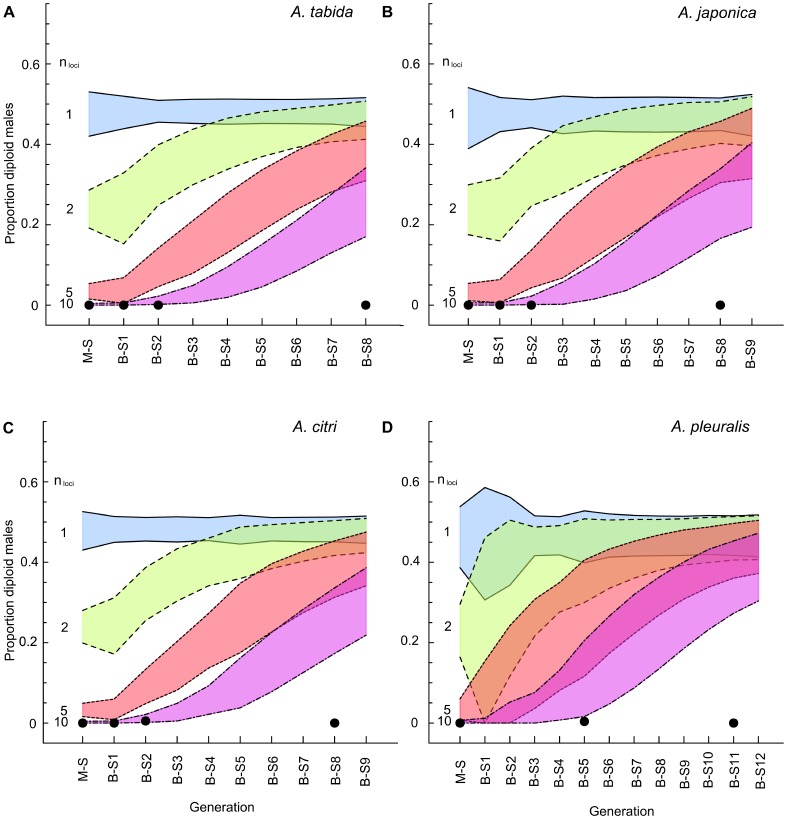
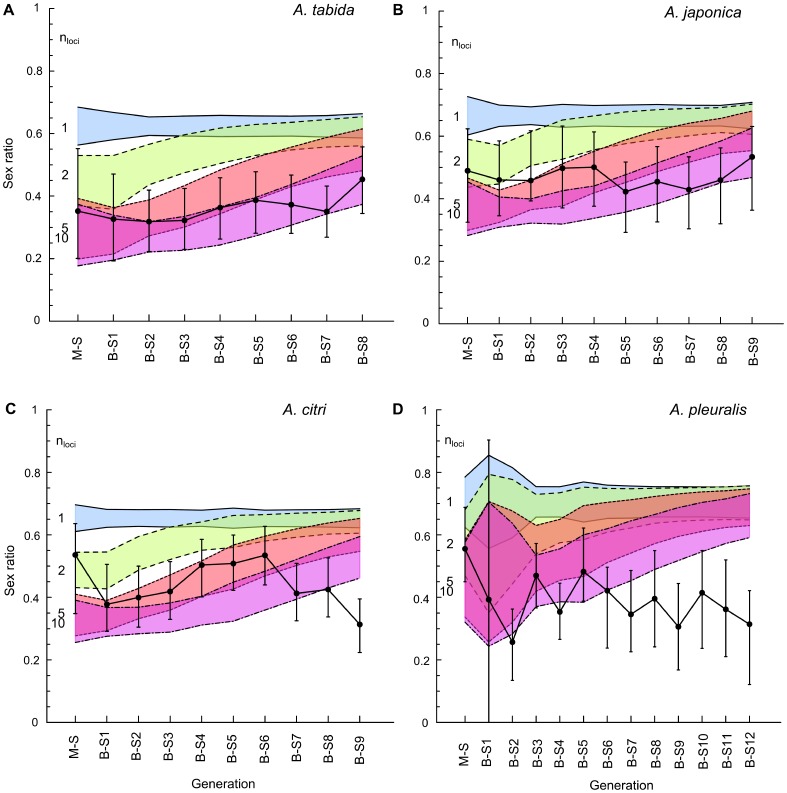
An attractive way to improve our understanding of sex determination evolution is to study the underlying mechanisms in closely related species and in a phylogenetic perspective. Hymenopterans are well suited owing to the diverse sex determination mechanisms, including different types of Complementary Sex Determination (CSD) and maternal control sex determination. We investigated different types of CSD in four species within the braconid wasp genus Asobara that exhibit diverse life-history traits. Nine to thirteen generations of inbreeding were monitored for diploid male production, brood size, offspring sex ratio, and pupal mortality as indicators for CSD. In addition, simulation models were developed to compare these observations to predicted patterns for multilocus CSD with up to ten loci. The inbreeding regime did not result in diploid male production, decreased brood sizes, substantially increased offspring sex ratios nor in increased pupal mortality. The simulations further allowed us to reject CSD with up to ten loci, which is a strong refutation of the multilocus CSD model. We discuss how the absence of CSD can be reconciled with the variation in life-history traits among Asobara species, and the ramifications for the phylogenetic distribution of sex determination mechanisms in the Hymenoptera.
Conflict of interest statement
Figures
Similar articles
-
Absence of single-locus complementary sex determination in the braconid wasps Asobara tabida and Alysia manducator.Heredity (Edinb). 2000 Jan;84 ( Pt 1):29-36. doi: 10.1046/j.1365-2540.2000.00628.x. Heredity (Edinb). 2000. PMID: 10692008
-
Absence of complementary sex determination in two Leptopilina species (Figitidae, Hymenoptera) and a reconsideration of its incompatibility with endosymbiont-induced thelytoky.Insect Sci. 2022 Jun;29(3):900-914. doi: 10.1111/1744-7917.12969. Epub 2021 Oct 25. Insect Sci. 2022. PMID: 34525260 Free PMC article.
-
Experimental support for multiple-locus complementary sex determination in the parasitoid Cotesia vestalis.Genetics. 2008 Nov;180(3):1525-35. doi: 10.1534/genetics.107.083907. Epub 2008 Sep 14. Genetics. 2008. PMID: 18791258 Free PMC article.
-
Sex determination in the hymenoptera.Annu Rev Entomol. 2008;53:209-30. doi: 10.1146/annurev.ento.53.103106.093441. Annu Rev Entomol. 2008. PMID: 17803453 Review.
-
Thelytoky and sex determination in the hymenoptera: mutual constraints.Sex Dev. 2014;8(1-3):50-8. doi: 10.1159/000356508. Epub 2013 Dec 7. Sex Dev. 2014. PMID: 24335186 Review.
Cited by
-
Genetics of decayed sexual traits in a parasitoid wasp with endosymbiont-induced asexuality.Heredity (Edinb). 2014 Nov;113(5):424-31. doi: 10.1038/hdy.2014.43. Epub 2014 Apr 30. Heredity (Edinb). 2014. PMID: 24781809 Free PMC article.
-
Quality of the Exotic Parasitoid Cotesia flavipes (Hymenoptera: Braconidae) Does Not Show Deleterious Effects after Inbreeding for 10 Generations.PLoS One. 2016 Aug 10;11(8):e0160898. doi: 10.1371/journal.pone.0160898. eCollection 2016. PLoS One. 2016. PMID: 27509087 Free PMC article.
-
A new biological species in the Mercurialis annua polyploid complex: functional divergence in inflorescence morphology and hybrid sterility.Ann Bot. 2019 Aug 2;124(1):165-178. doi: 10.1093/aob/mcz058. Ann Bot. 2019. PMID: 31098610 Free PMC article.
-
Bacterial Communities Differ among Drosophila melanogaster Populations and Affect Host Resistance against Parasitoids.PLoS One. 2016 Dec 14;11(12):e0167726. doi: 10.1371/journal.pone.0167726. eCollection 2016. PLoS One. 2016. PMID: 27973604 Free PMC article.
-
Natural and Artificial Selection for Parasitoid Resistance in Drosophila melanogaster Leave Different Genetic Signatures.Front Genet. 2019 May 31;10:479. doi: 10.3389/fgene.2019.00479. eCollection 2019. Front Genet. 2019. PMID: 31214243 Free PMC article.
References
-
- Kraak SBM, Pen I (2002) Sex-determining mechanisms in vertebrates. In: Hardy ICW editor. Sex ratios: Concepts and Research Methods. Cambridge: Cambridge University Press. 158–177.
-
- Saccone G, Pane A, Polito LC (2002) Sex determination in flies, fruitflies and butterflies. Genetica 116: 15–23. - PubMed
-
- Sánchez L (2008) Sex-determining mechanisms in insects. Int J Dev Biol 52: 837–856. - PubMed
-
- Bull JJ (1985) Sex determining mechanisms: an evolutionary perspective. Experientia 41: 1285–1289. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
