

Microgravity induces pelvic bone loss through osteoclastic activity, osteocytic osteolysis, and osteoblastic cell cycle inhibition by CDKN1a/p21
- PMID: 23637819
- PMCID: PMC3630201
- DOI: 10.1371/journal.pone.0061372
Microgravity induces pelvic bone loss through osteoclastic activity, osteocytic osteolysis, and osteoblastic cell cycle inhibition by CDKN1a/p21
Abstract
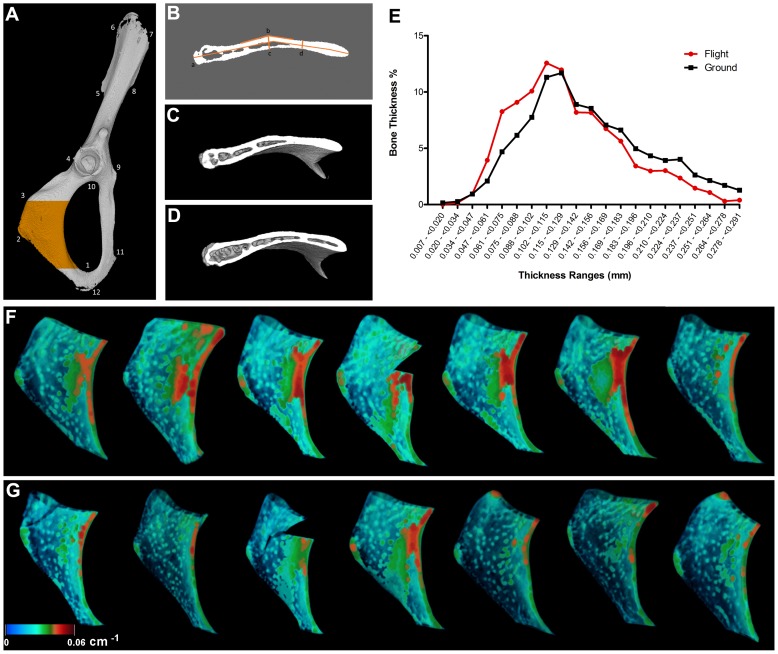
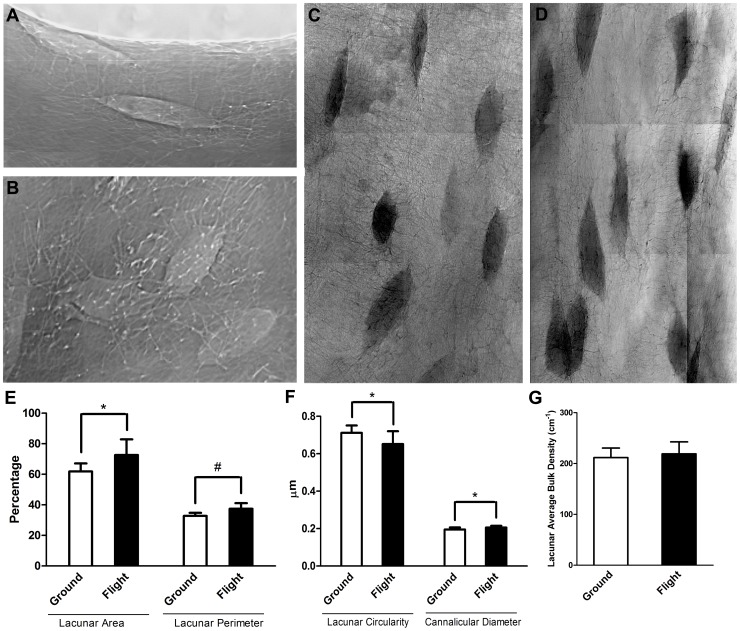
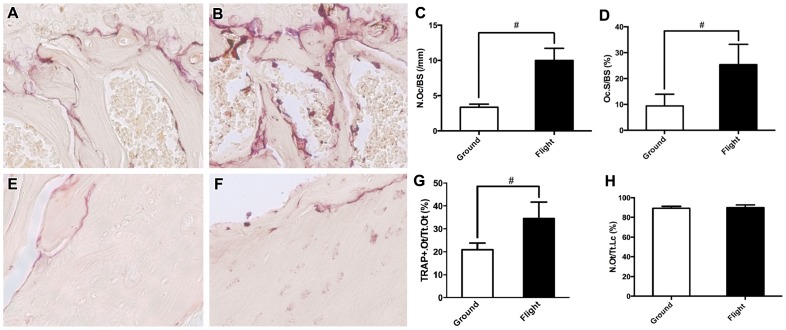
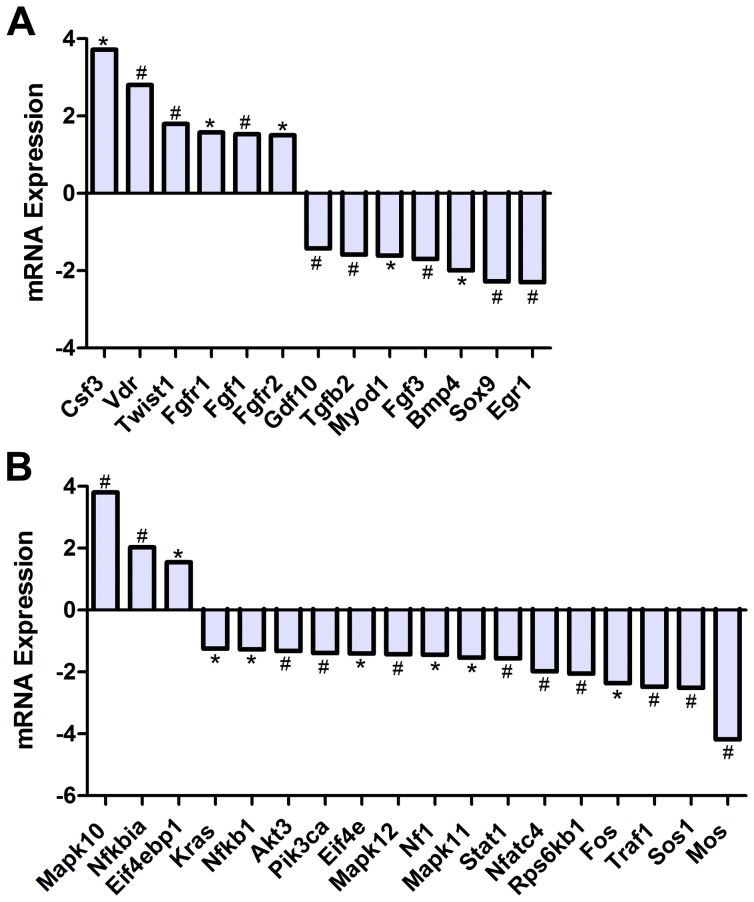
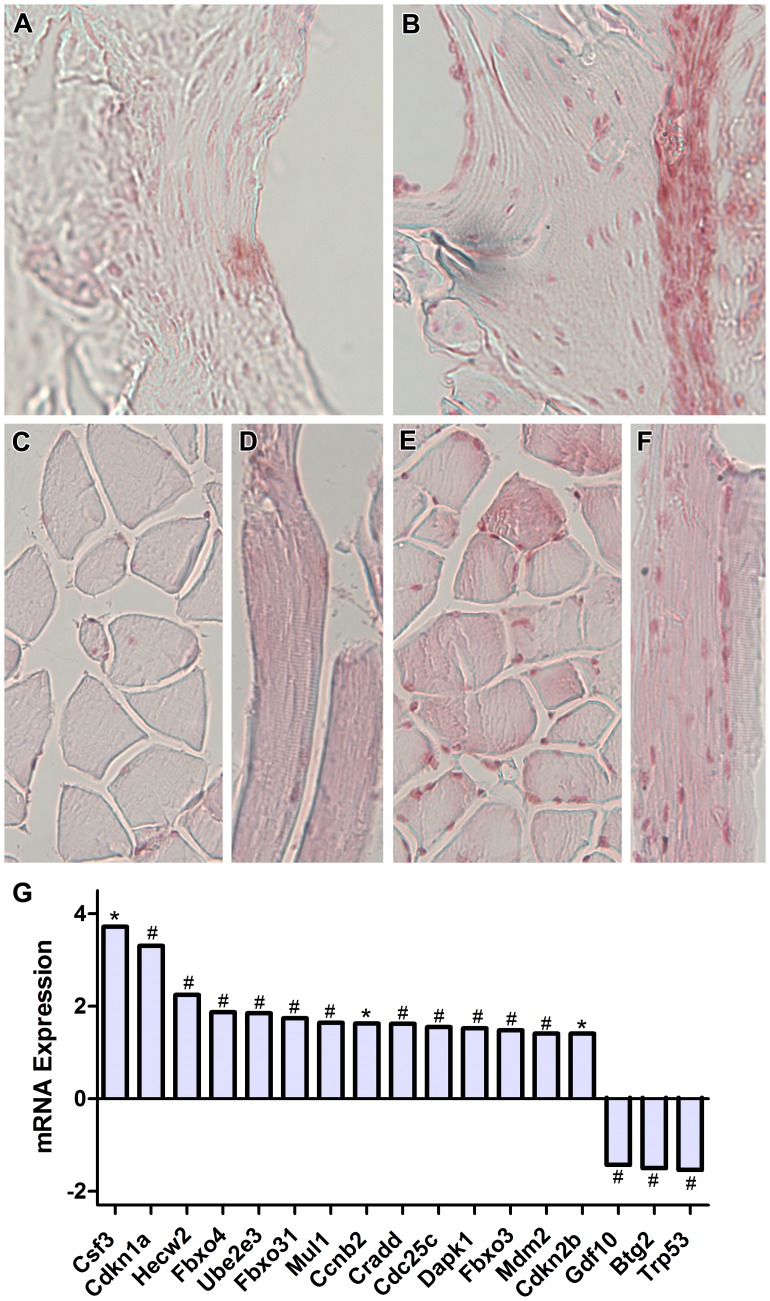
Bone is a dynamically remodeled tissue that requires gravity-mediated mechanical stimulation for maintenance of mineral content and structure. Homeostasis in bone occurs through a balance in the activities and signaling of osteoclasts, osteoblasts, and osteocytes, as well as proliferation and differentiation of their stem cell progenitors. Microgravity and unloading are known to cause osteoclast-mediated bone resorption; however, we hypothesize that osteocytic osteolysis, and cell cycle arrest during osteogenesis may also contribute to bone loss in space. To test this possibility, we exposed 16-week-old female C57BL/6J mice (n = 8) to microgravity for 15-days on the STS-131 space shuttle mission. Analysis of the pelvis by µCT shows decreases in bone volume fraction (BV/TV) of 6.29%, and bone thickness of 11.91%. TRAP-positive osteoclast-covered trabecular bone surfaces also increased in microgravity by 170% (p = 0.004), indicating osteoclastic bone degeneration. High-resolution X-ray nanoCT studies revealed signs of lacunar osteolysis, including increases in cross-sectional area (+17%, p = 0.022), perimeter (+14%, p = 0.008), and canalicular diameter (+6%, p = 0.037). Expression of matrix metalloproteinases (MMP) 1, 3, and 10 in bone, as measured by RT-qPCR, was also up-regulated in microgravity (+12.94, +2.98 and +16.85 fold respectively, p<0.01), with MMP10 localized to osteocytes, and consistent with induction of osteocytic osteolysis. Furthermore, expression of CDKN1a/p21 in bone increased 3.31 fold (p<0.01), and was localized to osteoblasts, possibly inhibiting the cell cycle during tissue regeneration as well as conferring apoptosis resistance to these cells. Finally the apoptosis inducer Trp53 was down-regulated by -1.54 fold (p<0.01), possibly associated with the quiescent survival-promoting function of CDKN1a/p21. In conclusion, our findings identify the pelvic and femoral region of the mouse skeleton as an active site of rapid bone loss in microgravity, and indicate that this loss is not limited to osteoclastic degradation. Therefore, this study offers new evidence for microgravity-induced osteocytic osteolysis, and CDKN1a/p21-mediated osteogenic cell cycle arrest.
Conflict of interest statement
Figures

References
-
- Nabavi N, Khandani A, Camirand A, Harrison RE (2011) Effects of microgravity on osteoclast bone resorption and osteoblast cytoskeletal organization and adhesion. Bone 49: 965–974. - PubMed
-
- Hughes-Fulford M (2003) Function of the cytoskeleton in gravisensing during spaceflight. Adv Space Res 32: 1585–1593. - PubMed
-
- Ohshima H (2006) [Bone loss and bone metabolism in astronauts during long-duration space flight]. Clin Calcium 16: 81–85. - PubMed
-
- Smith SM, Heer M (2002) Calcium and bone metabolism during space flight. Nutrition 18: 849–852. - PubMed
-
- Zayzafoon M, Meyers VE, McDonald JM (2005) Microgravity: the immune response and bone. Immunol Rev 208: 267–280. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
