

Ultrastructure of protrusive actin filament arrays
- PMID: 23639311
- PMCID: PMC3758367
- DOI: 10.1016/j.ceb.2013.04.003
Ultrastructure of protrusive actin filament arrays
Abstract
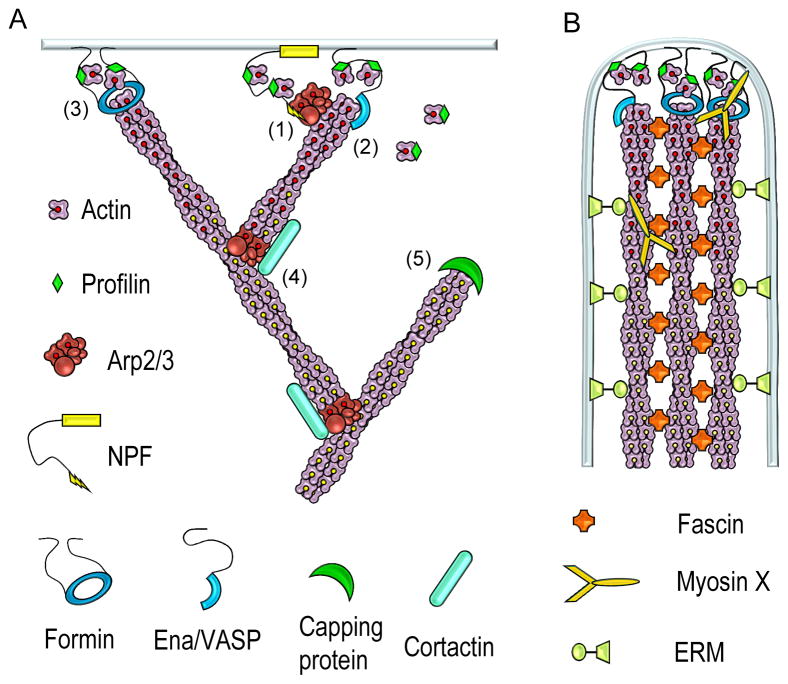
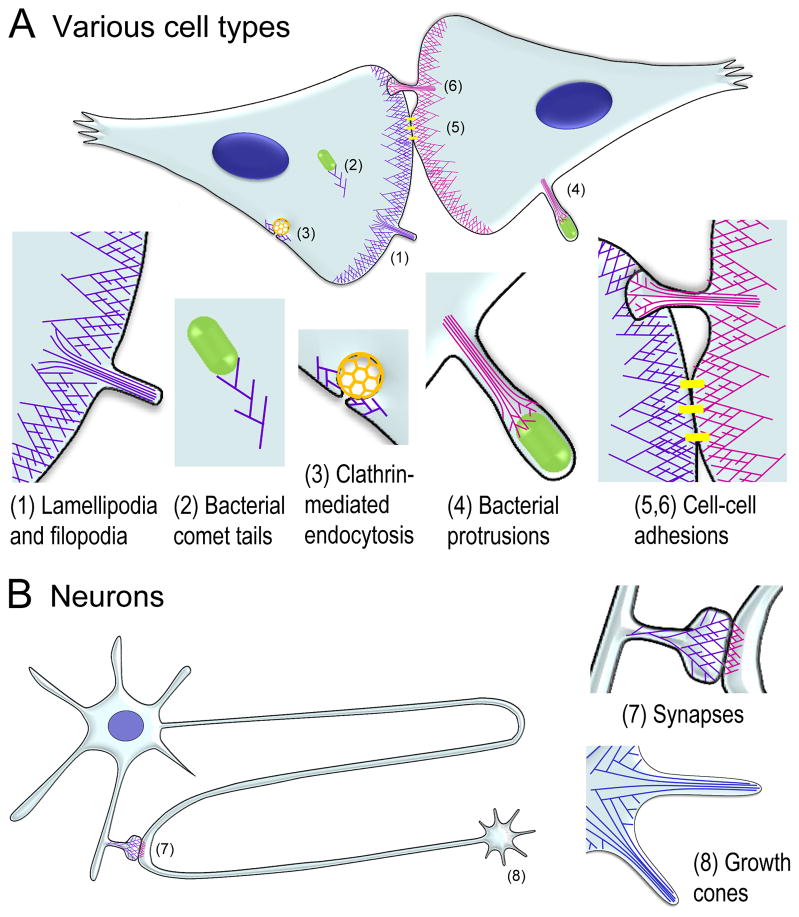
The actin cytoskeleton is the major force-generating machinery in the cell, which can produce pushing, pulling, and resistance forces. To accomplish these diverse functions, actin filaments, with help of numerous accessory proteins, form higher order ensembles, networks and bundles, adapted to specific tasks. Moreover, dynamic properties of the actin cytoskeleton allow a cell to constantly build, renew, and redesign actin structures according to its changing needs. High resolution architecture of actin filament arrays provides key information for understanding mechanisms of force generation. To generate pushing force, cells use coordinated polymerization of multiple actin filaments organized into branched (dendritic) networks or parallel bundles. This review summarizes our current knowledge of the structural organization of these two actin filament arrays.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Ultrastructure of the actin cytoskeleton.Curr Opin Cell Biol. 2018 Oct;54:1-8. doi: 10.1016/j.ceb.2018.02.007. Epub 2018 Feb 21. Curr Opin Cell Biol. 2018. PMID: 29477121 Free PMC article. Review.
-
Actin filament organization within dendrites and dendritic spines during development.Brain Res. 1986 Jun;392(1-2):263-9. doi: 10.1016/0165-3806(86)90253-1. Brain Res. 1986. PMID: 3708380
-
The Actin Cytoskeleton and Actin-Based Motility.Cold Spring Harb Perspect Biol. 2018 Jan 2;10(1):a018267. doi: 10.1101/cshperspect.a018267. Cold Spring Harb Perspect Biol. 2018. PMID: 29295889 Free PMC article. Review.
-
Interactions of ADF/cofilin, Arp2/3 complex, capping protein and profilin in remodeling of branched actin filament networks.Curr Biol. 2000 Oct 19;10(20):1273-82. doi: 10.1016/s0960-9822(00)00749-1. Curr Biol. 2000. PMID: 11069108
-
Probing actin polymerization by intermolecular cross-linking.J Cell Biol. 1988 Mar;106(3):785-96. doi: 10.1083/jcb.106.3.785. J Cell Biol. 1988. PMID: 3346326 Free PMC article.
Cited by
-
Actin cytoskeleton in mesenchymal-to-amoeboid transition of cancer cells.Int Rev Cell Mol Biol. 2020;356:197-256. doi: 10.1016/bs.ircmb.2020.06.002. Epub 2020 Jul 16. Int Rev Cell Mol Biol. 2020. PMID: 33066874 Free PMC article. Review.
-
Two Motors and One Spring: Hypothetic Roles of Non-Muscle Myosin II and Submembrane Actin-Based Cytoskeleton in Cell Volume Sensing.Int J Mol Sci. 2021 Jul 26;22(15):7967. doi: 10.3390/ijms22157967. Int J Mol Sci. 2021. PMID: 34360739 Free PMC article. Review.
-
Arp2/3 Complex Is Required for Macrophage Integrin Functions but Is Dispensable for FcR Phagocytosis and In Vivo Motility.Dev Cell. 2017 Sep 11;42(5):498-513.e6. doi: 10.1016/j.devcel.2017.08.003. Epub 2017 Aug 31. Dev Cell. 2017. PMID: 28867487 Free PMC article.
-
In vitro model of bone to facilitate measurement of adhesion forces and super-resolution imaging of osteoclasts.Sci Rep. 2016 Mar 3;6:22585. doi: 10.1038/srep22585. Sci Rep. 2016. PMID: 26935172 Free PMC article.
-
Pimozide suppresses cancer cell migration and tumor metastasis through binding to ARPC2, a subunit of the Arp2/3 complex.Cancer Sci. 2019 Dec;110(12):3788-3801. doi: 10.1111/cas.14205. Epub 2019 Nov 15. Cancer Sci. 2019. PMID: 31571309 Free PMC article.
References
-
- Small JV, Isenberg G, Celis JE. Polarity of actin at the leading edge of cultured cells. Nature. 1978;272:638–639. - PubMed
-
- Svitkina TM, Shevelev AA, Bershadsky AD, Gelfand VI. Cytoskeleton of mouse embryo fibroblasts. Electron microscopy of platinum replicas. Eur J Cell Biol. 1984;34:64–74. - PubMed
-
- Ben-Harush K, Maimon T, Patla I, Villa E, Medalia O. Visualizing cellular processes at the molecular level by cryo-electron tomography. J Cell Sci. 2010;123:7–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
