

E. coli ClpA catalyzed polypeptide translocation is allosterically controlled by the protease ClpP
- PMID: 23639359
- PMCID: PMC3918674
- DOI: 10.1016/j.jmb.2013.04.019
E. coli ClpA catalyzed polypeptide translocation is allosterically controlled by the protease ClpP
Abstract
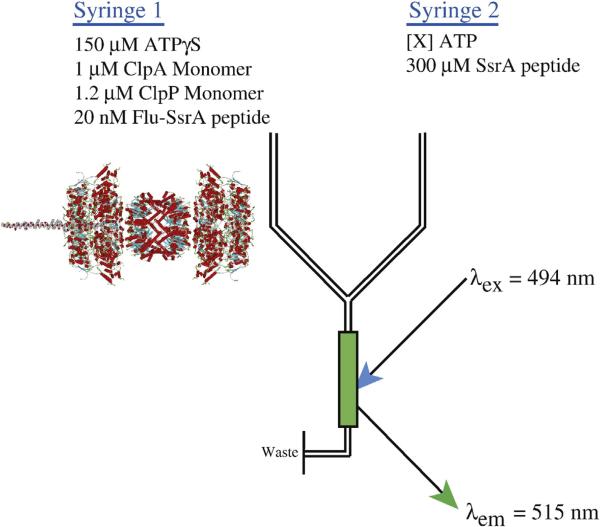
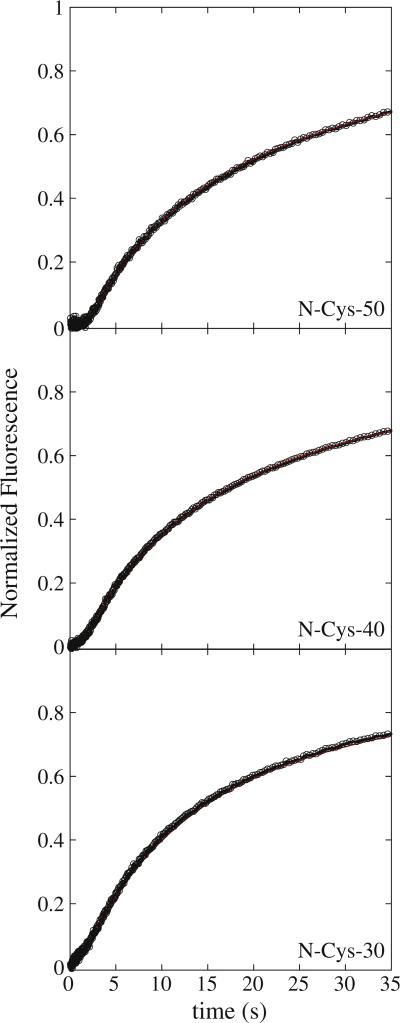
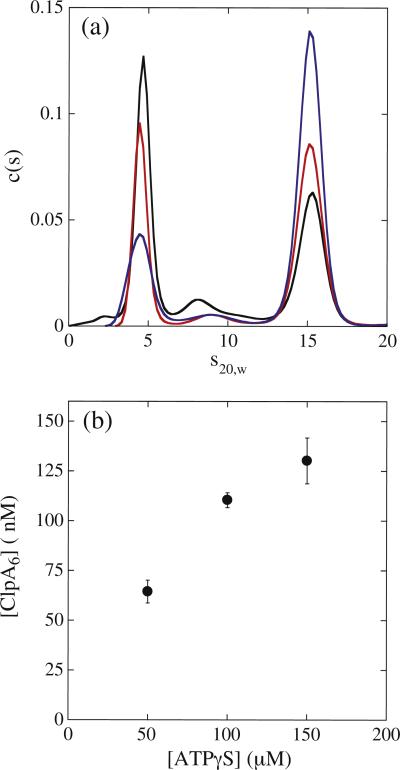
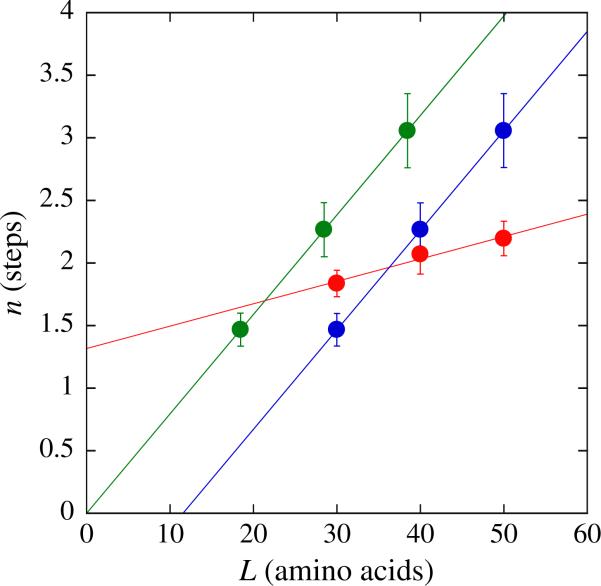
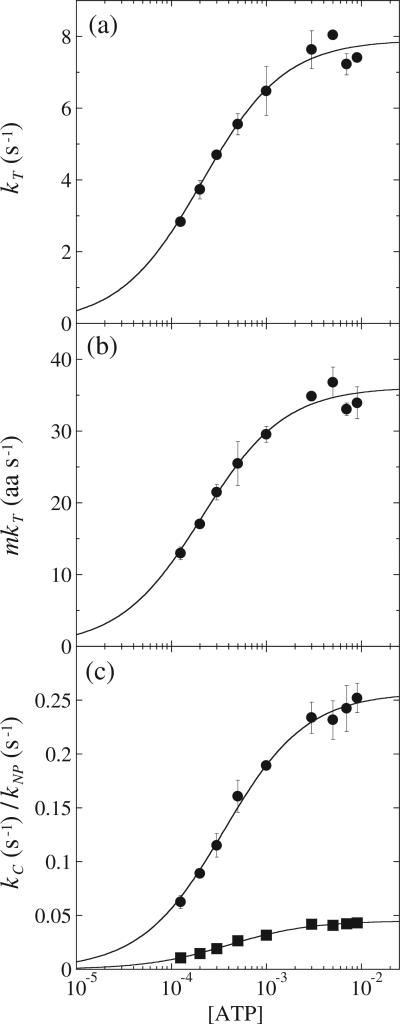
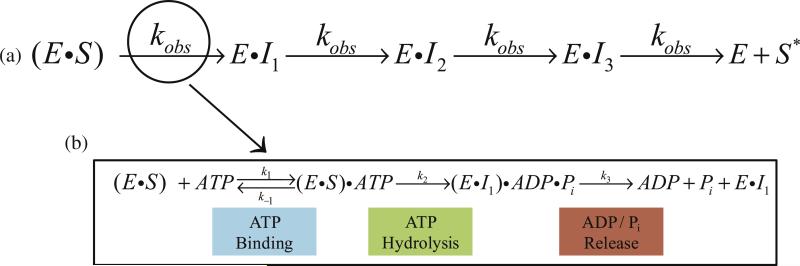
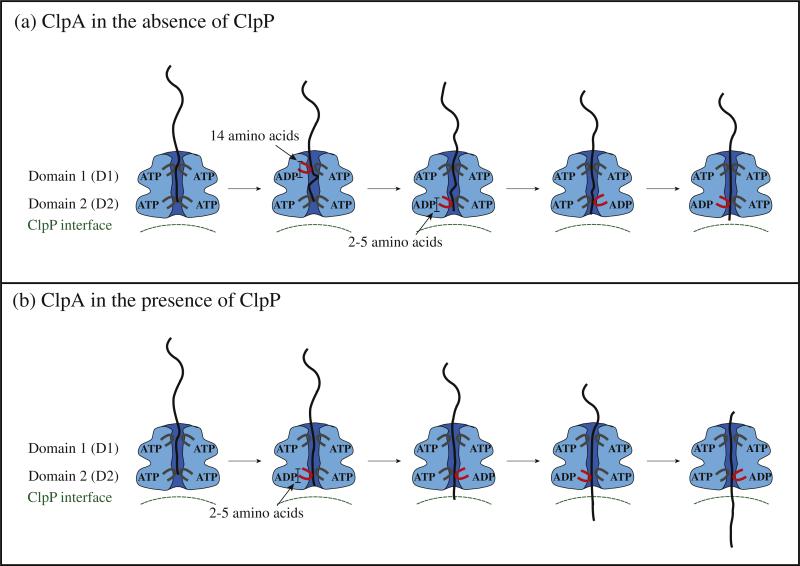
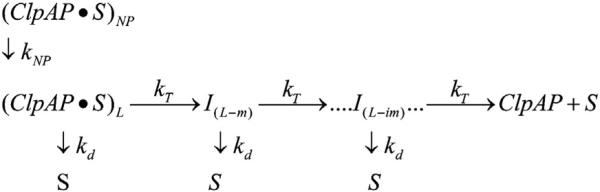
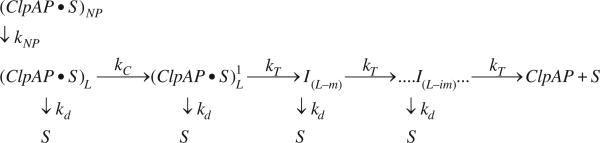
There are five known ATP-dependent proteases in Escherichia coli (Lon, ClpAP, ClpXP, HslUV, and the membrane-associated FtsH) that catalyze the removal of both misfolded and properly folded proteins in cellular protein quality control pathways. Hexameric ClpA rings associate with one or both faces of the cylindrically shaped tetradecameric ClpP protease. ClpA catalyzes unfolding and translocation of polypeptide substrates into the proteolytic core of ClpP for degradation through repeated cycles of ATP binding and hydrolysis at two nucleotide binding domains on each ClpA monomer. We previously reported a molecular mechanism for ClpA catalyzed polypeptide translocation in the absence of ClpP, including elementary rate constants, overall rate, and the kinetic step size. However, the potential allosteric effect of ClpP on the mechanism of ClpA catalyzed translocation remains unclear. Using single-turnover fluorescence stopped-flow methods, here we report that ClpA, when associated with ClpP, translocates polypeptide with an overall rate of ~35 aa s(-1) and, on average, traverses ~5 aa between two rate-limiting steps with reduced cooperativity between ATP binding sites in the hexameric ring. This is in direct contrast to our previously reported observation that, in the absence of ClpP, ClpA translocates polypeptide substrates with a maximum translocation rate of ~20 aa s(-1) with cooperativity between ATPase sites. Our results demonstrate that ClpP allosterically impacts the polypeptide translocation activity of ClpA by reducing the cooperativity between ATP binding sites.
Keywords: AAA+ motor proteins; ATP-dependent proteases; ATPγS; EDTA; FRET; GFP; NLLS; adenosine 5′-(γ-thio)-triphosphate; ethylenediaminetetraacetic acid; fluorescence resonance energy transfer; green fluorescent protein; nonlinear least squares; pre-steady-state kinetics; protein unfoldases.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Alberts B. The cell as a collection of protein machines: preparing the next generation of molecular biologists. Cell. 1998;92:291–294. - PubMed
-
- Neuwald AF, Aravind L, Spouge JL, Koonin EV. AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999;9:27–43. - PubMed
-
- Gottesman S. Proteases and their targets in Escherichia coli. Annu. Rev. Genet. 1996;30:465–506. - PubMed
-
- Sauer RT, Baker TA. AAA+ proteases: ATP-fueled machines of protein destruction. Annu. Rev. Biochem. 2011;80:587–612. - PubMed
-
- Maglica Z, Kolygo K, Weber-Ban E. Optimal efficiency of ClpAP and ClpXP chaperone-proteases is achieved by architectural symmetry. Structure. 2009;17:508–516. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
