

The neuropilin 1 cytoplasmic domain is required for VEGF-A-dependent arteriogenesis
- PMID: 23639442
- PMCID: PMC3774154
- DOI: 10.1016/j.devcel.2013.03.019
The neuropilin 1 cytoplasmic domain is required for VEGF-A-dependent arteriogenesis
Abstract
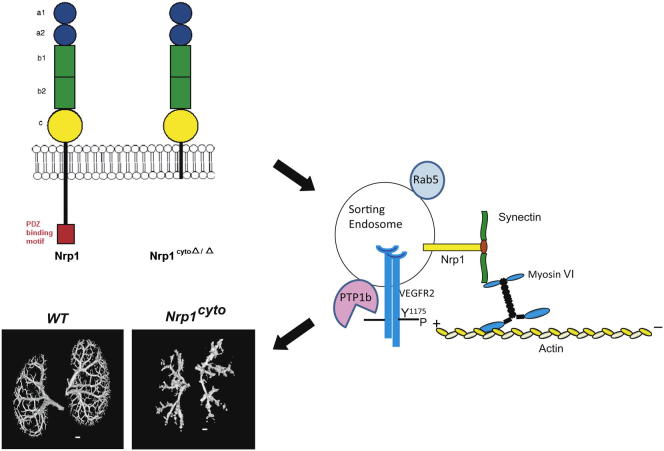
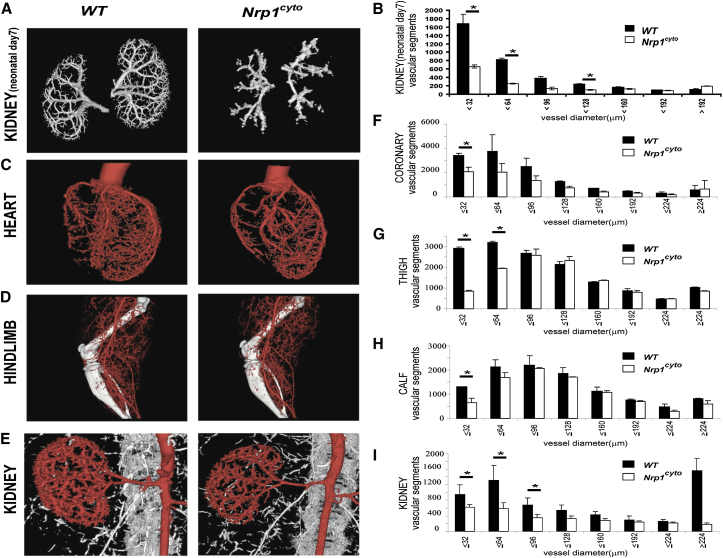
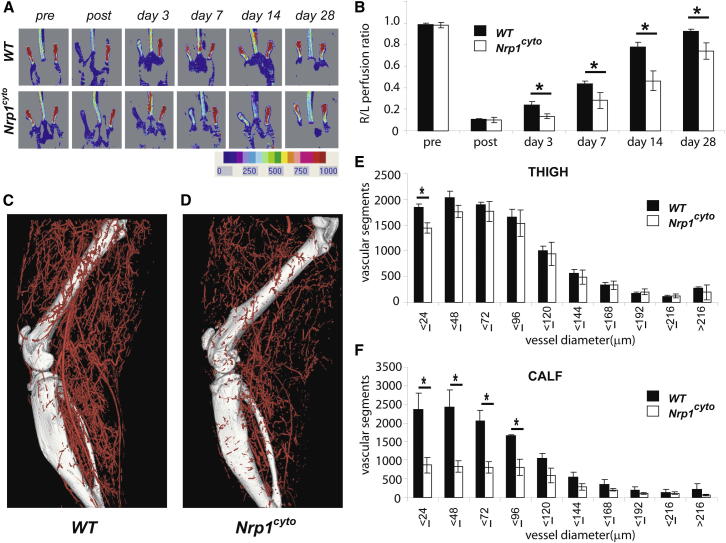
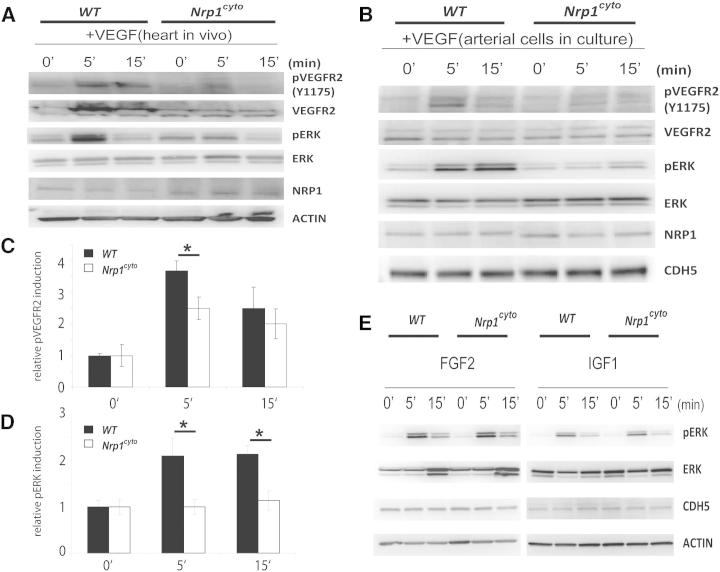
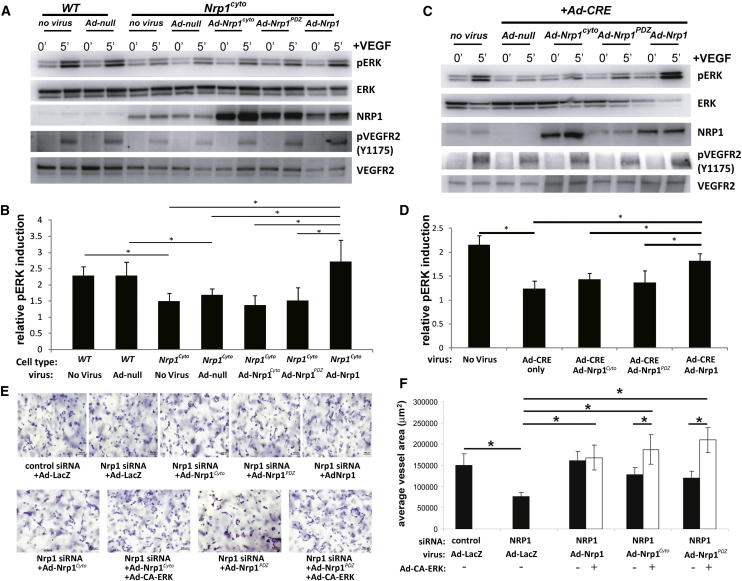
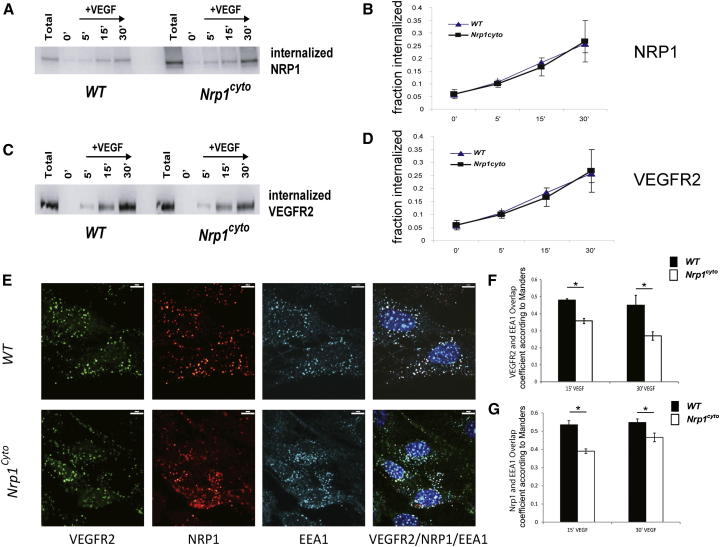
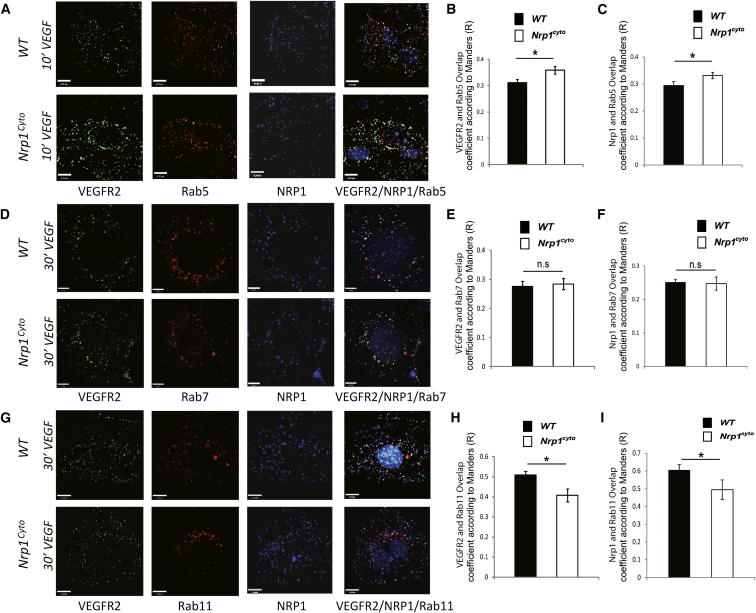
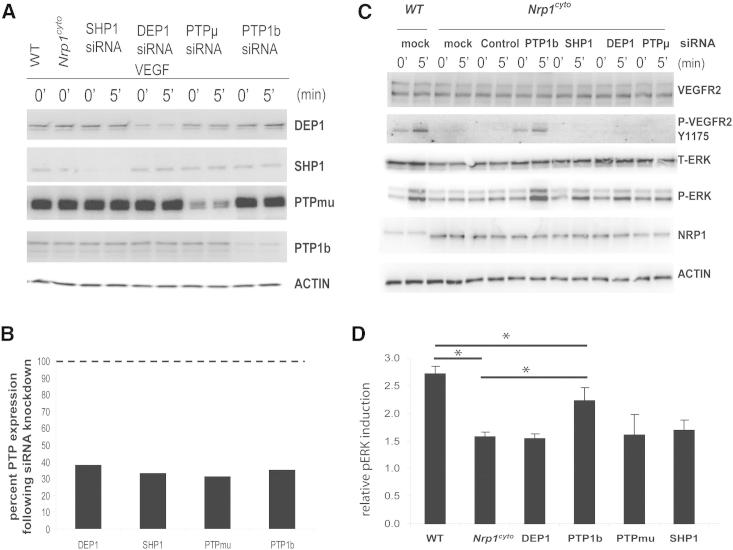
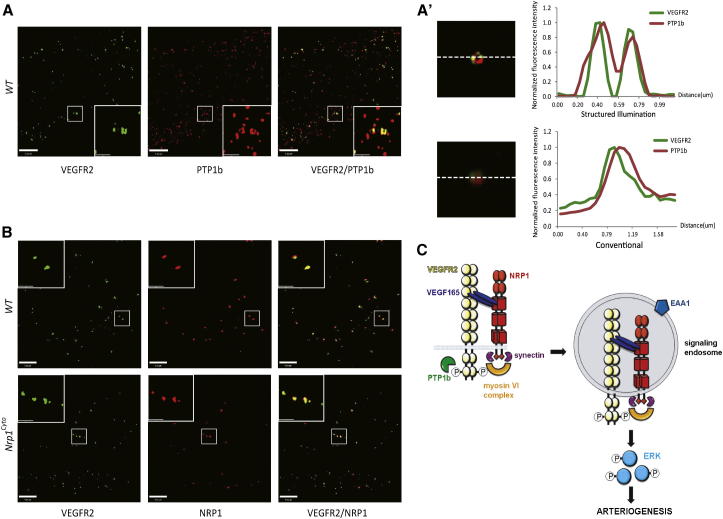
Neuropilin 1 (NRP1) plays an important but ill-defined role in VEGF-A signaling and vascular morphogenesis. We show that mice with a knockin mutation that ablates the NRP1 cytoplasmic tail (Nrp1(cyto)) have normal angiogenesis but impaired developmental and adult arteriogenesis. The arteriogenic defect was traced to the absence of a PDZ-dependent interaction between NRP1 and VEGF receptor 2 (VEGFR2) complex and synectin, which delayed trafficking of endocytosed VEGFR2 from Rab5+ to EAA1+ endosomes. This led to increased PTPN1 (PTP1b)-mediated dephosphorylation of VEGFR2 at Y(1175), the site involved in activating ERK signaling. The Nrp1(cyto) mutation also impaired endothelial tubulogenesis in vitro, which could be rescued by expressing full-length NRP1 or constitutively active ERK. These results demonstrate that the NRP1 cytoplasmic domain promotes VEGFR2 trafficking in a PDZ-dependent manner to regulate arteriogenic ERK signaling and establish a role for NRP1 in VEGF-A signaling during vascular morphogenesis.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Ballmer-Hofer K., Andersson A.E., Ratcliffe L.E., Berger P. Neuropilin-1 promotes VEGFR-2 trafficking through Rab11 vesicles thereby specifying signal output. Blood. 2011;118:816–826. - PubMed
-
- Booth R.A., Cummings C., Tiberi M., Liu X.J. GIPC participates in G protein signaling downstream of insulin-like growth factor 1 receptor. J. Biol. Chem. 2002;277:6719–6725. - PubMed
-
- Chittenden T.W., Claes F., Lanahan A.A., Autiero M., Palac R.T., Tkachenko E.V., Elfenbein A., Ruiz de Almodovar C., Dedkov E., Tomanek R. Selective regulation of arterial branching morphogenesis by synectin. Dev. Cell. 2006;10:783–795. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
