

Nitric-oxide synthase knockout modulates Ca²⁺-sensing receptor expression and signaling in mouse mesenteric arteries
- PMID: 23639802
- PMCID: PMC3684838
- DOI: 10.1124/jpet.113.205534
Nitric-oxide synthase knockout modulates Ca²⁺-sensing receptor expression and signaling in mouse mesenteric arteries
Abstract
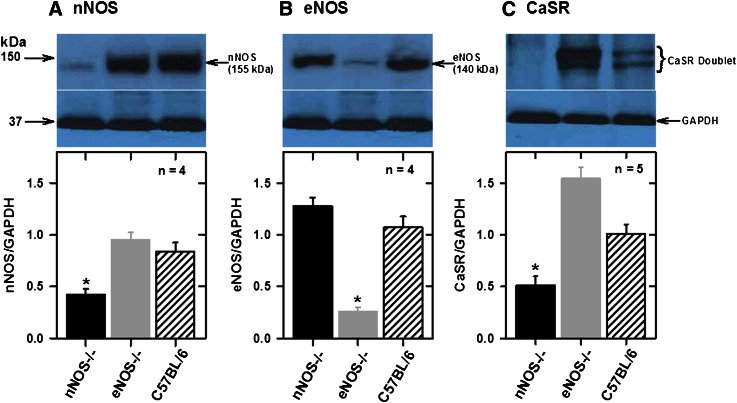
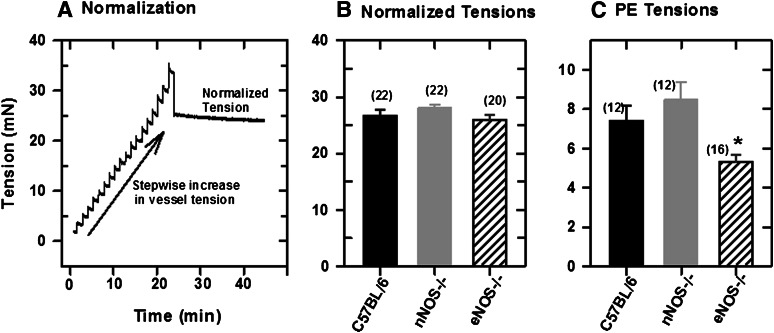
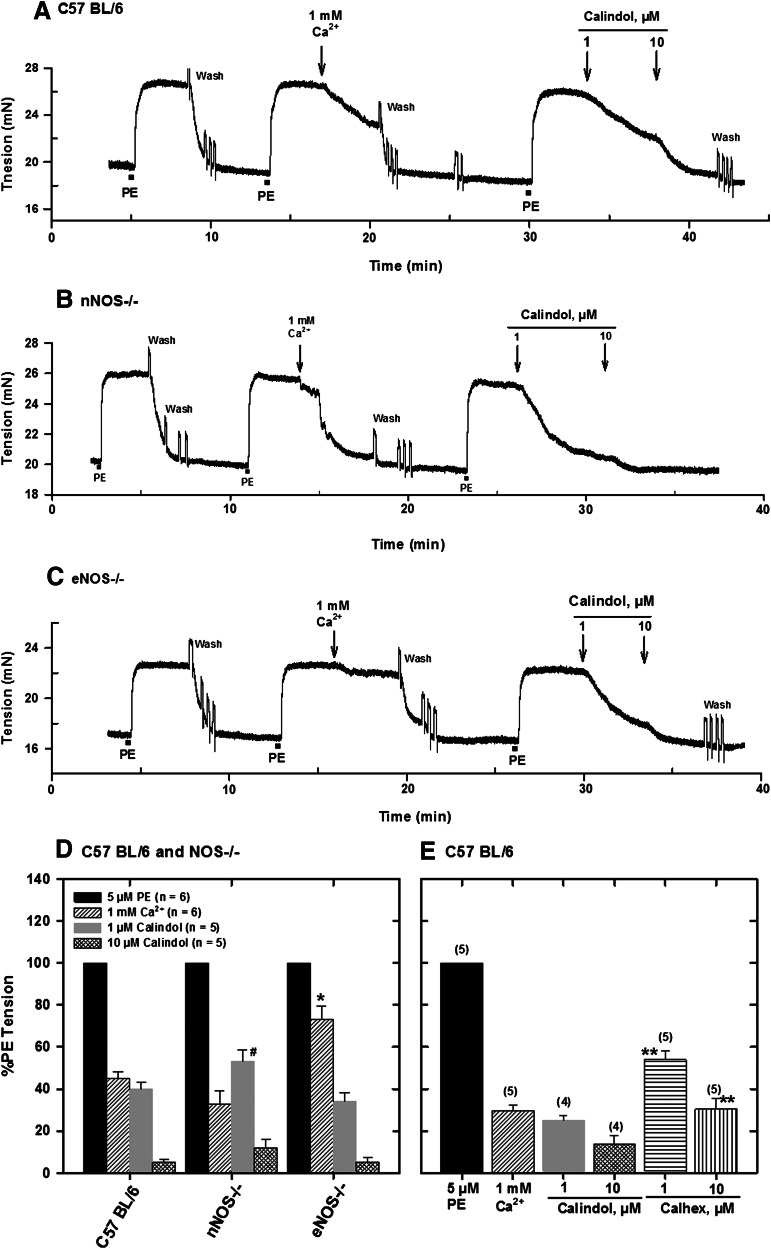
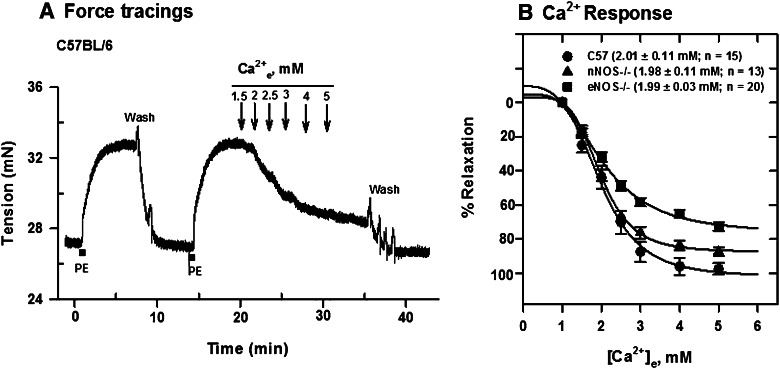
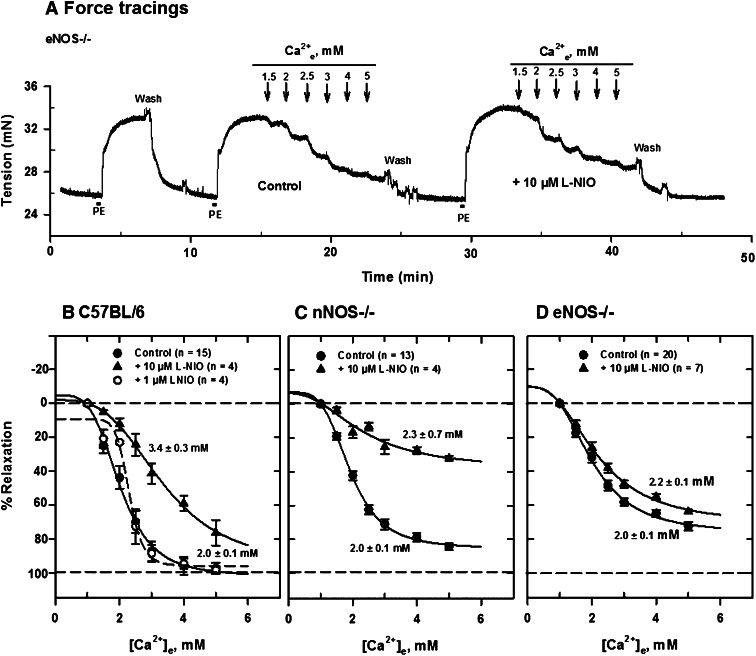
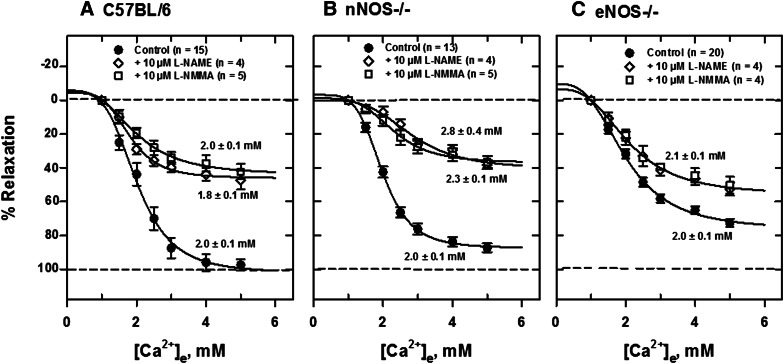
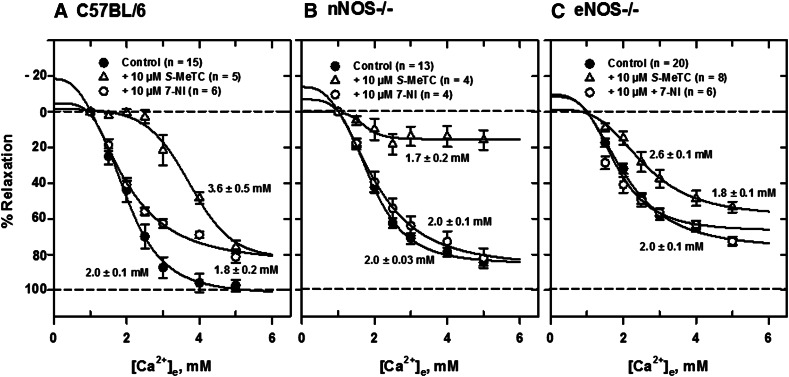
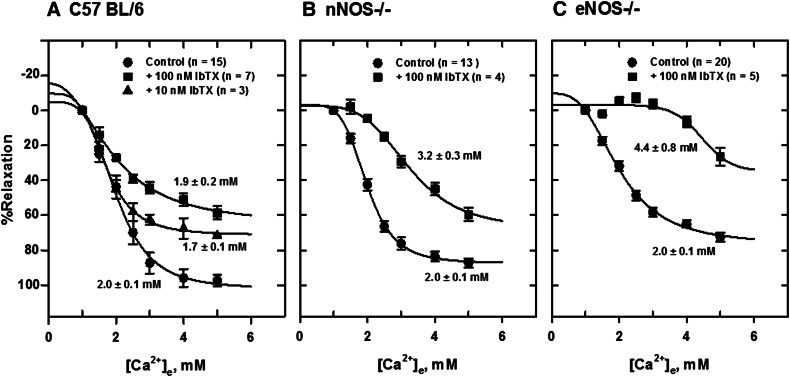
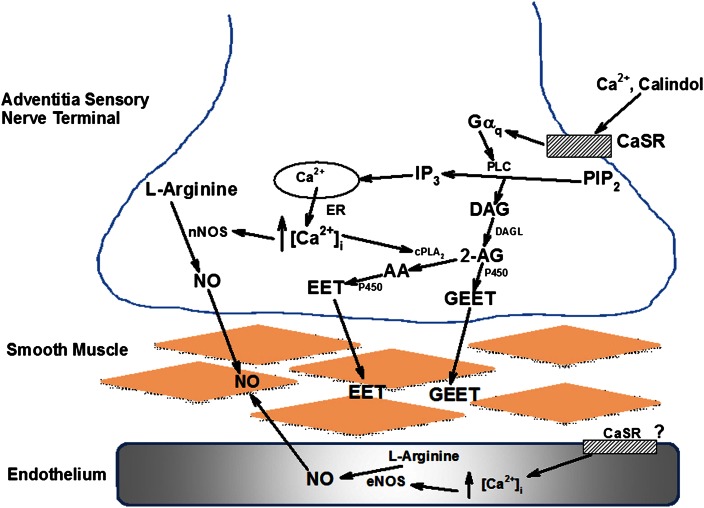
Extracellular calcium (Ca²⁺(e))-induced relaxation of isolated, phenylephrine (PE)-contracted mesenteric arteries is dependent on an intact perivascular sensory nerve network that expresses the Ca²⁺-sensing receptor (CaSR). Activation of the receptor stimulates an endocannabinoid vasodilator pathway, which is dependent on cytochrome P450 and phospholipase A₂ but largely independent of the endothelium. In the present study, we determined the role of nitric oxide (NO) in perivascular nerve CaSR-mediated relaxation of PE-contracted mesenteric resistance arteries isolated from mice. Using automated wire myography, we studied the effects of NO synthase (NOS) gene knockout (NOS(-/-)) and pharmacologic inhibition of NOS on Ca²⁺(e)-induced relaxation of PE-contracted arteries. Endothelial NOS knockout (eNOS(-/-)) upregulates but neuronal NOS knockout (nNOS(-/-)) downregulates CaSR expression. NOS(-/-) reduced maximum Ca²⁺(e)-induced relaxation with no change in EC₅₀ values, with eNOS(-/-) having the largest effect. The responses of vessels to calindol and Calhex 231 indicate that the CaSR mediates relaxation. L-N⁵-(1-iminoethyl)-ornithine reduced Ca²⁺(e)-induced relaxation of PE-contracted arteries from C57BL/6 control mice by ≈38% but had a smaller effect in vessels from eNOS(-/-) mice. 7-Nitroindazole had no significant effect on relaxation of arteries from NOS(-/-) mice, but both N(G)-nitro-L-arginine methylester and N(G)-monomethyl-L-arginine significantly reduced the relaxation maxima in all groups. Interestingly, the nNOS-selective inhibitor S-methyl-L-thiocitrulline significantly increased the EC₅₀ value by ≈60% in tissues from C57BL/6 mice but reduced the maximum response by ≈80% in those from nNOS(-/-) mice. Ca²⁺-activated big potassium channels play a major role in the process, as demonstrated by the effect of iberiotoxin. We conclude that CaSR signaling in mesenteric arteries stimulates eNOS and NO production that regulates Ca²⁺(e)-induced relaxation.
Figures
References
-
- Alam MU, Kirton JP, Wilkinson FL, Towers E, Sinha S, Rouhi M, Vizard TN, Sage AP, Martin D, Ward DT, et al. (2009) Calcification is associated with loss of functional calcium-sensing receptor in vascular smooth muscle cells. Cardiovasc Res 81:260–268 - PubMed
-
- Angus JA, Wright CE. (2000) Techniques to study the pharmacodynamics of isolated large and small blood vessels. J Pharmacol Toxicol Methods 44:395–407 - PubMed
-
- Awumey EM, Howlett AC, Putney JW, Jr, Diz DI, Bukoski RD. (2007) Ca(2+) mobilization through dorsal root ganglion Ca(2+)-sensing receptor stably expressed in HEK293 cells. Am J Physiol Cell Physiol 292:C1895–C1905 - PubMed
-
- Bai M, Quinn S, Trivedi S, Kifor O, Pearce SH, Pollak MR, Krapcho K, Hebert SC, Brown EM. (1996) Expression and characterization of inactivating and activating mutations in the human Ca2+o-sensing receptor. J Biol Chem 271:19537–19545 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
