

Multisensory control of hippocampal spatiotemporal selectivity
- PMID: 23641063
- PMCID: PMC4049564
- DOI: 10.1126/science.1232655
Multisensory control of hippocampal spatiotemporal selectivity
Abstract
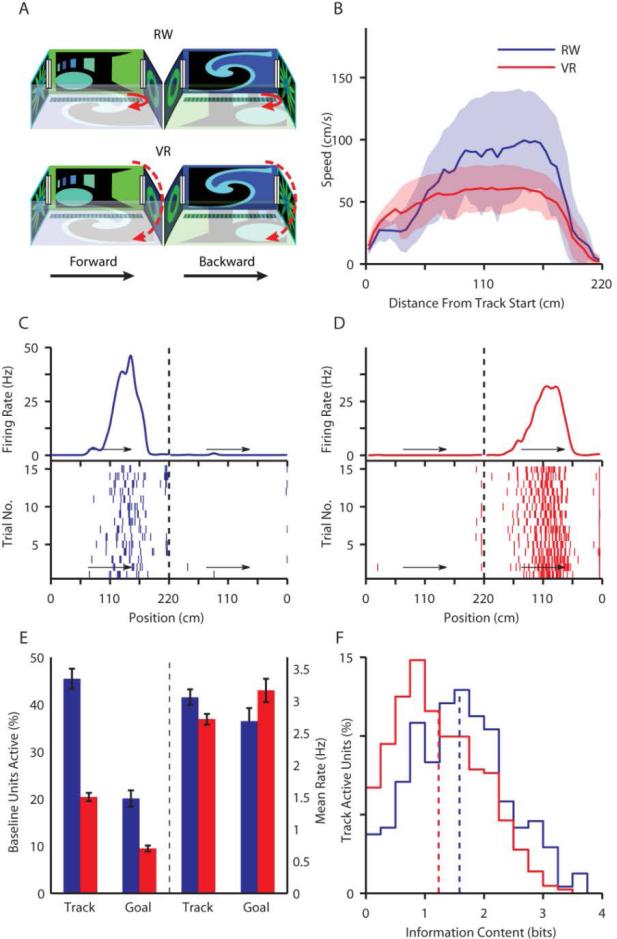
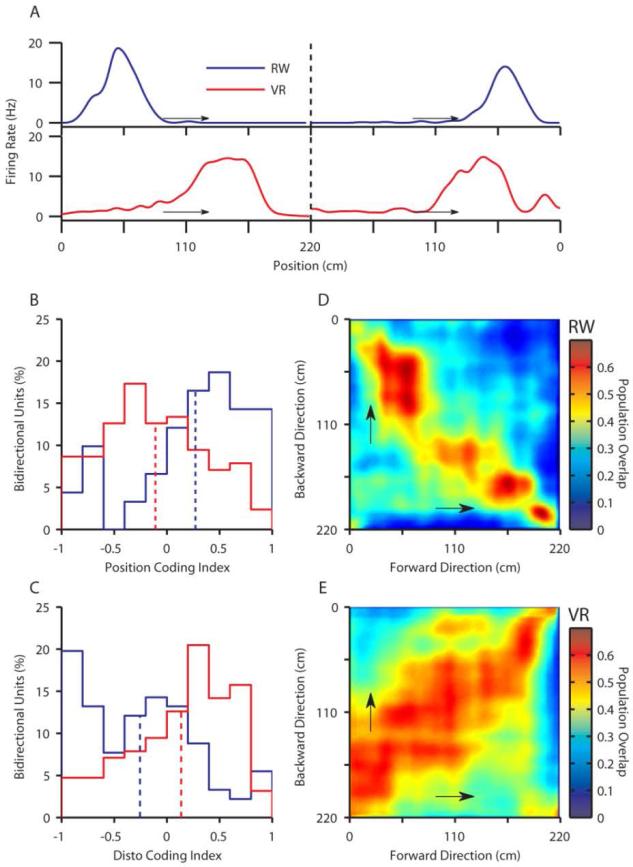
The hippocampal cognitive map is thought to be driven by distal visual cues and self-motion cues. However, other sensory cues also influence place cells. Hence, we measured rat hippocampal activity in virtual reality (VR), where only distal visual and nonvestibular self-motion cues provided spatial information, and in the real world (RW). In VR, place cells showed robust spatial selectivity; however, only 20% were track active, compared with 45% in the RW. This indicates that distal visual and nonvestibular self-motion cues are sufficient to provide selectivity, but vestibular and other sensory cues present in RW are necessary to fully activate the place-cell population. In addition, bidirectional cells preferentially encoded distance along the track in VR, while encoding absolute position in RW. Taken together, these results suggest the differential contributions of these sensory cues in shaping the hippocampal population code. Theta frequency was reduced, and its speed dependence was abolished in VR, but phase precession was unaffected, constraining mechanisms governing both hippocampal theta oscillations and temporal coding. These results reveal cooperative and competitive interactions between sensory cues for control over hippocampal spatiotemporal selectivity and theta rhythm.
Figures


References
-
- O'Keefe J, Dostrovsky J. The hippocampus as a spatial map: Preliminary evidence from unit activity in the freely-moving rat. Brain Research. 1971;34:171–175. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
