

Sugar metabolism and the plant target of rapamycin kinase: a sweet operaTOR?
- PMID: 23641244
- PMCID: PMC3640205
- DOI: 10.3389/fpls.2013.00093
Sugar metabolism and the plant target of rapamycin kinase: a sweet operaTOR?
Abstract
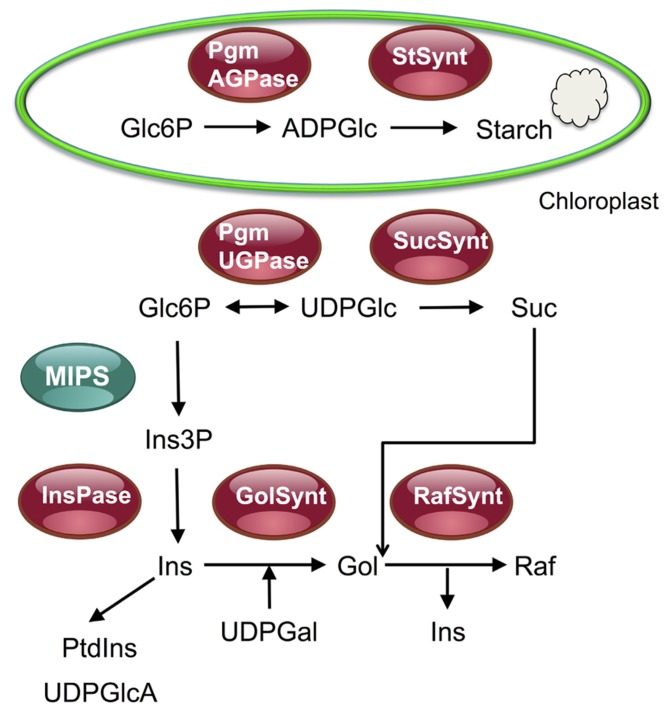
In eukaryotes, the ubiquitous TOR (target of rapamycin) kinase complexes have emerged as central regulators of cell growth and metabolism. The plant TOR complex 1 (TORC1), that contains evolutionary conserved protein partners, has been shown to be implicated in various aspects of C metabolism. Indeed Arabidopsis lines affected in the expression of TORC1 components show profound perturbations in the metabolism of several sugars, including sucrose, starch, and raffinose. Metabolite profiling experiments coupled to transcriptomic analyses of lines affected in TORC1 expression also reveal a wider deregulation of primary metabolism. Moreover recent data suggest that the kinase activity of TORC1, which controls biological outputs like mRNA translation or autophagy, is directly regulated by soluble sugars.
Keywords: TOR serine-threonine kinases; myo-inositol-1-phosphate synthase; raffinose; starch; target of rapamycin.
Figures

References
-
- Ball S., Colleoni C., Cenci U., Raj J. N., Tirtiaux C. (2011). The evolution of glycogen and starch metabolism in eukaryotes gives molecular clues to understand the establishment of plastid endosymbiosis. J. Exp. Bot. 62 1775–1801 - PubMed
-
- Binda M., Peli-Gulli M. P., Bonfils G., Panchaud N., Urban J., Sturgill T. W., et al. (2009). The Vam6 GEF controls TORC1 by activating the EGO complex. Mol. Cell 35 563–573 - PubMed
-
- Caldana C., Li Y., Leisse A., Zhang Y., Bartholomaeus L., Fernie A. R., et al. (2013). Systemic analysis of inducible target of rapamycin mutants reveal a general metabolic switch controlling growth in Arabidopsis thaliana. Plant J. 73 897–909 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases