

N-terminal truncation of an isolated human IgG1 CH2 domain significantly increases its stability and aggregation resistance
- PMID: 23641816
- PMCID: PMC3795862
- DOI: 10.1021/mp400075f
N-terminal truncation of an isolated human IgG1 CH2 domain significantly increases its stability and aggregation resistance
Abstract
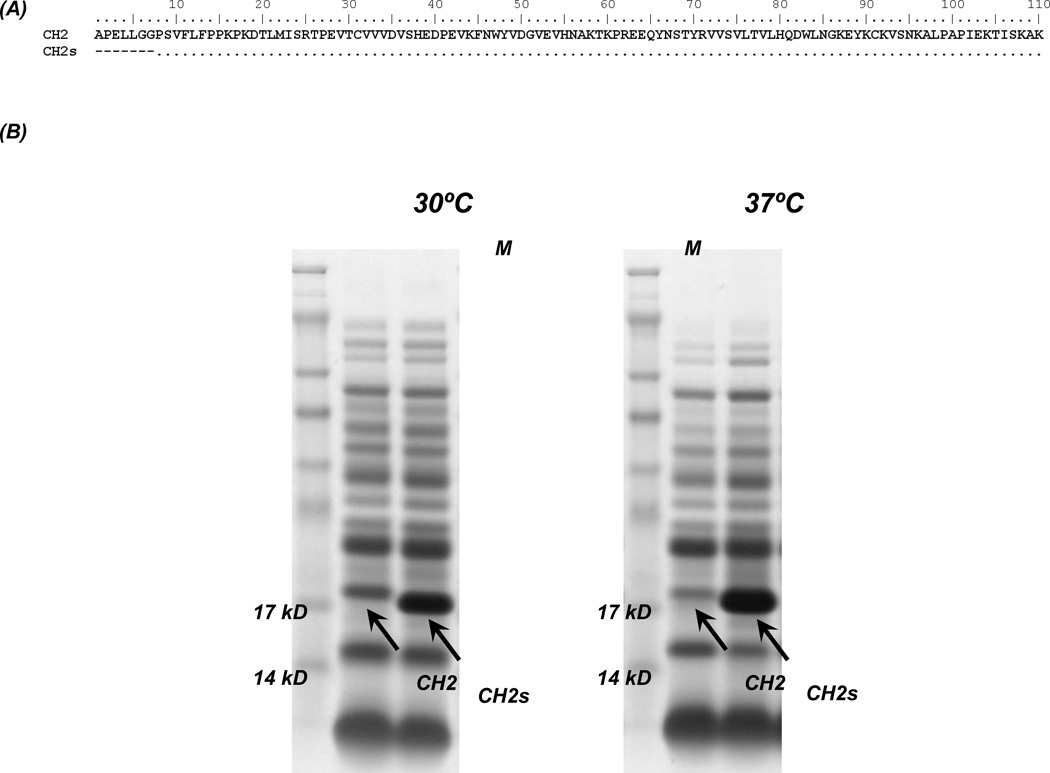
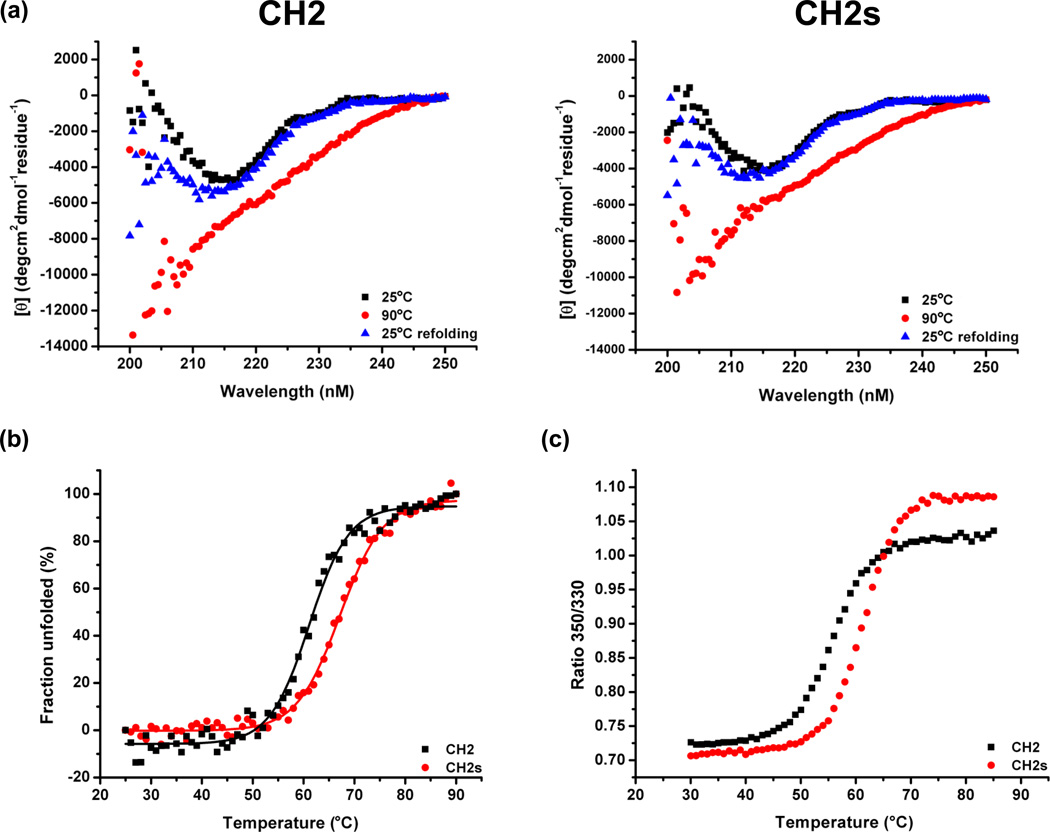
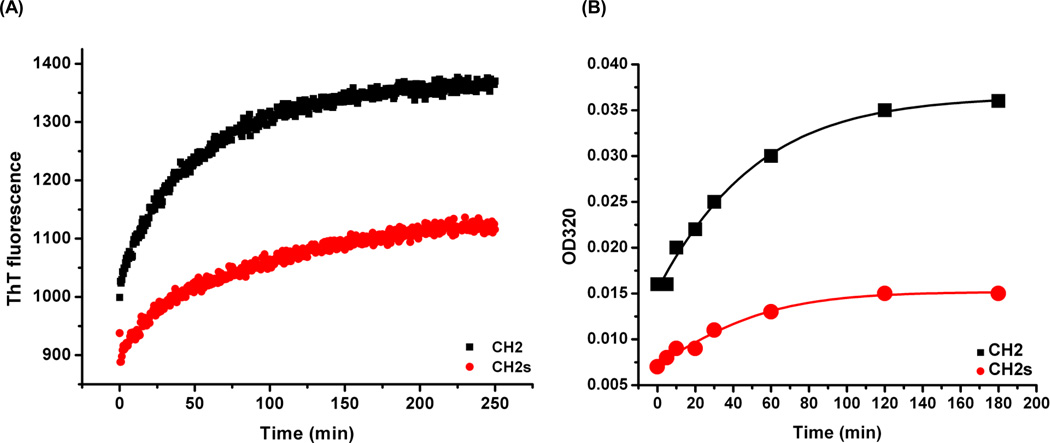
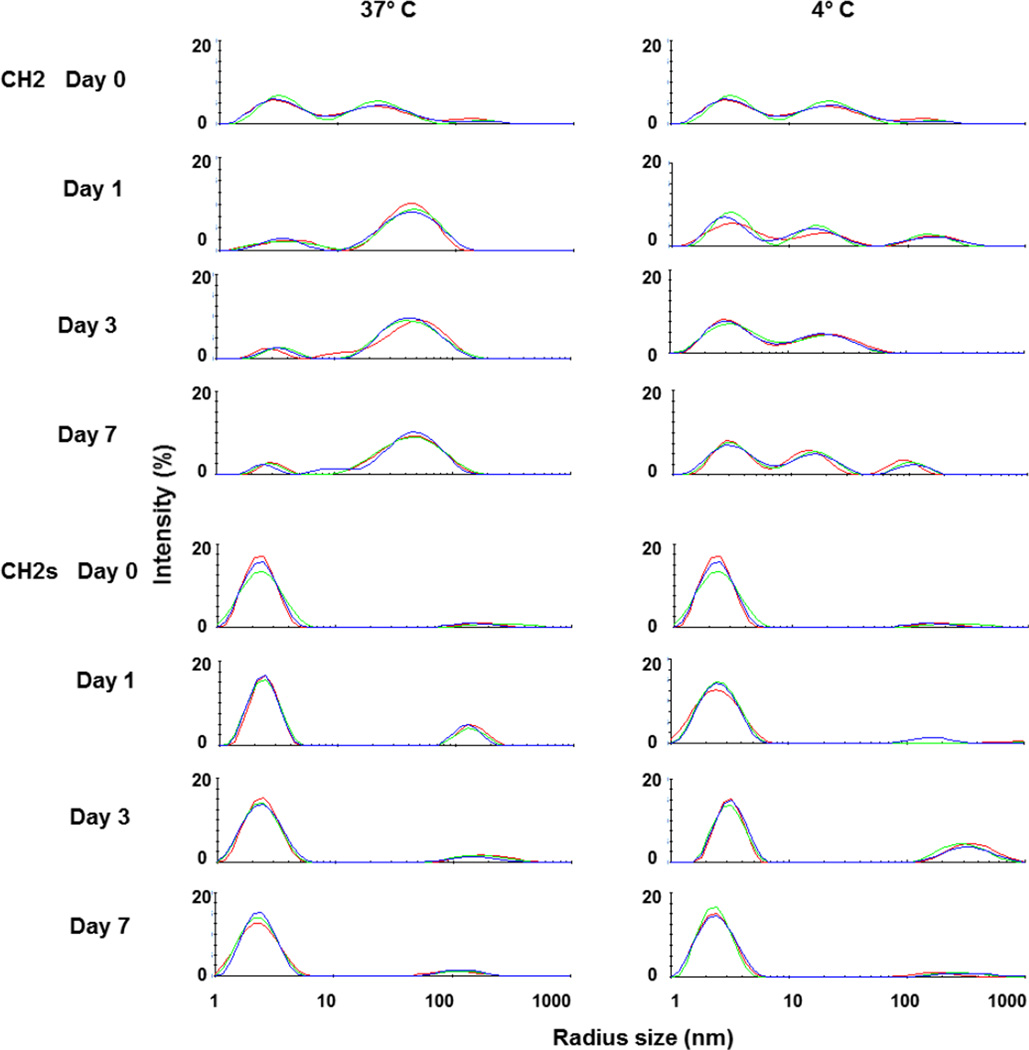
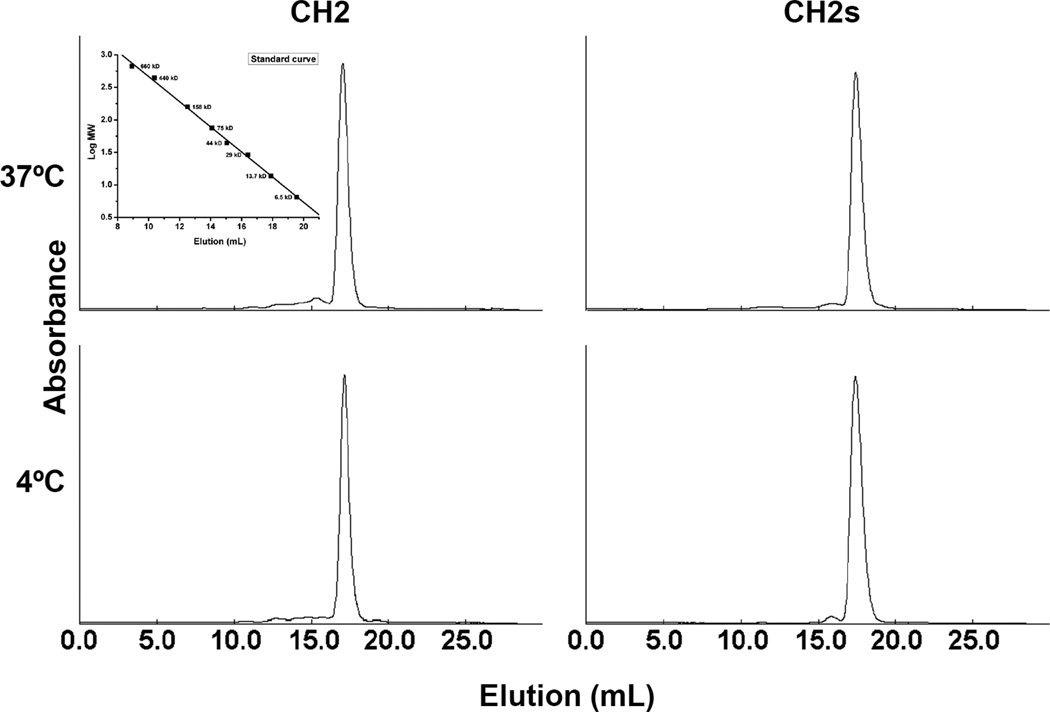
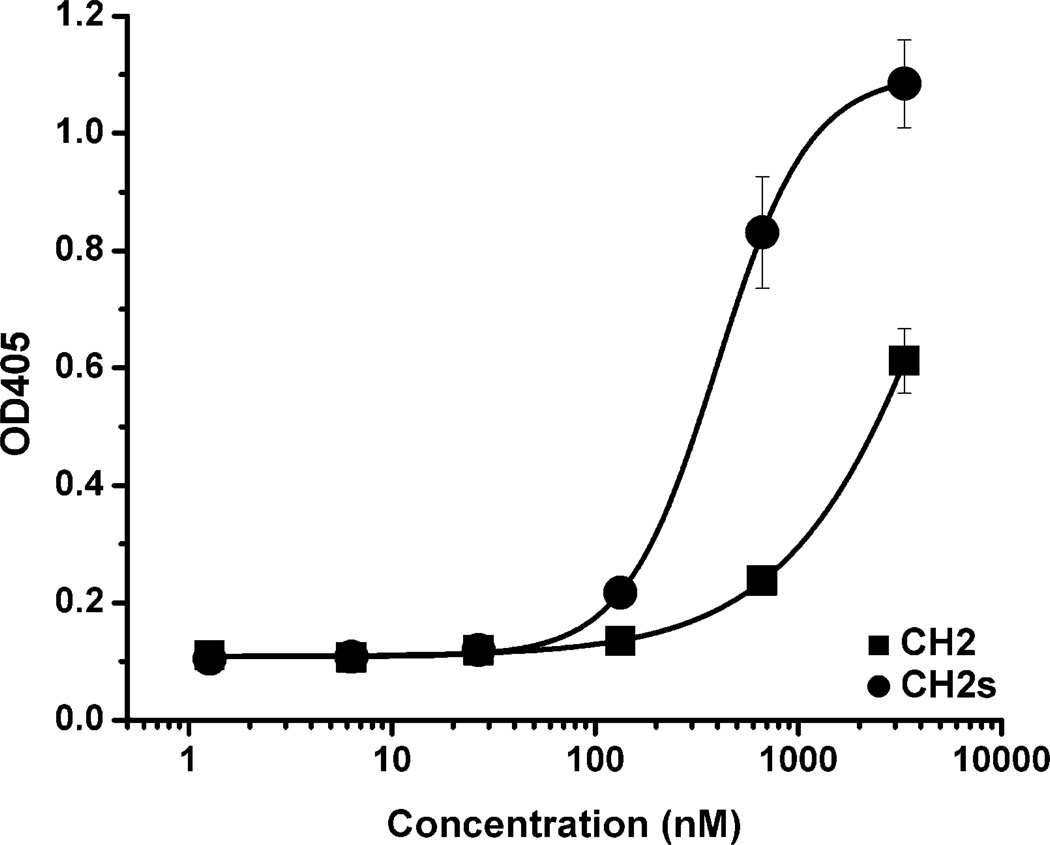
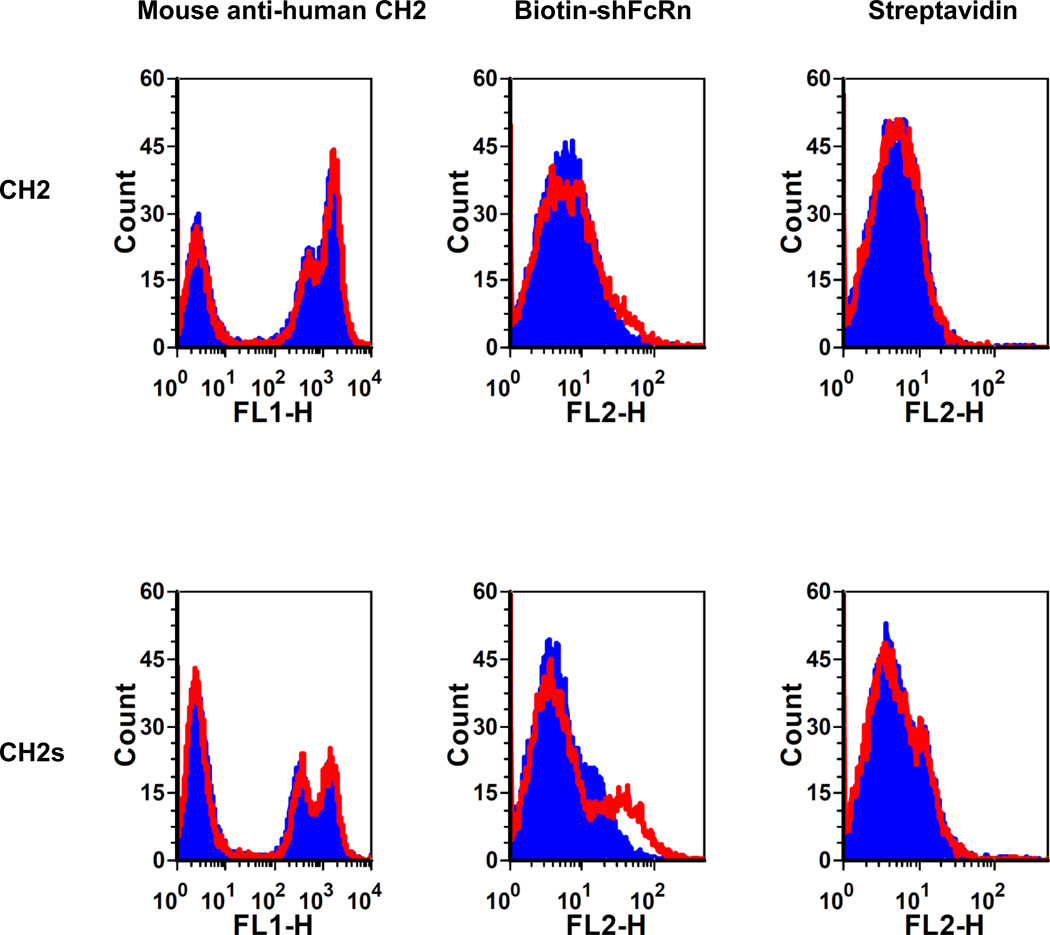
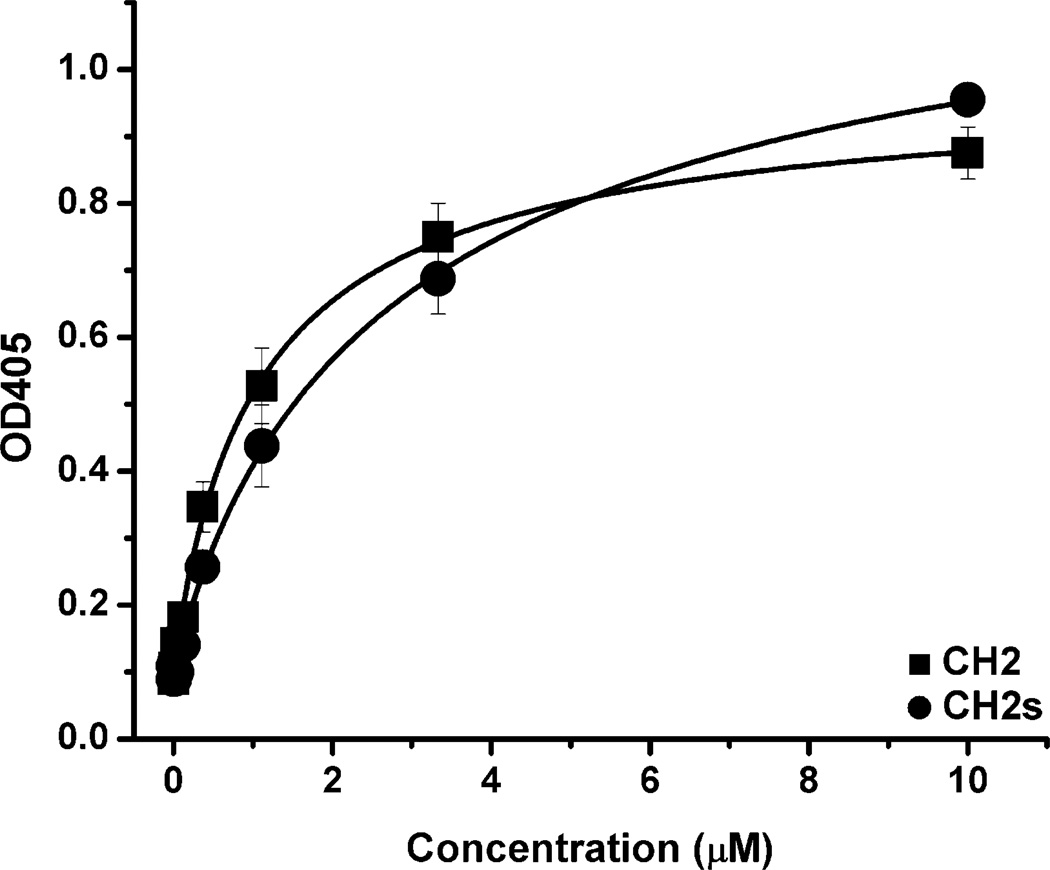
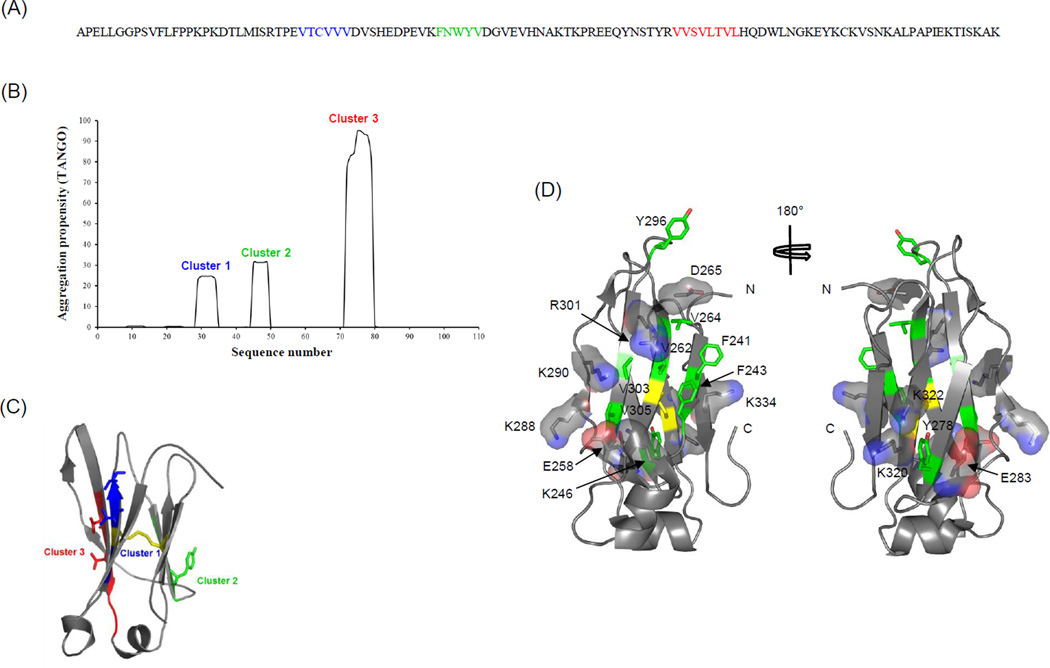
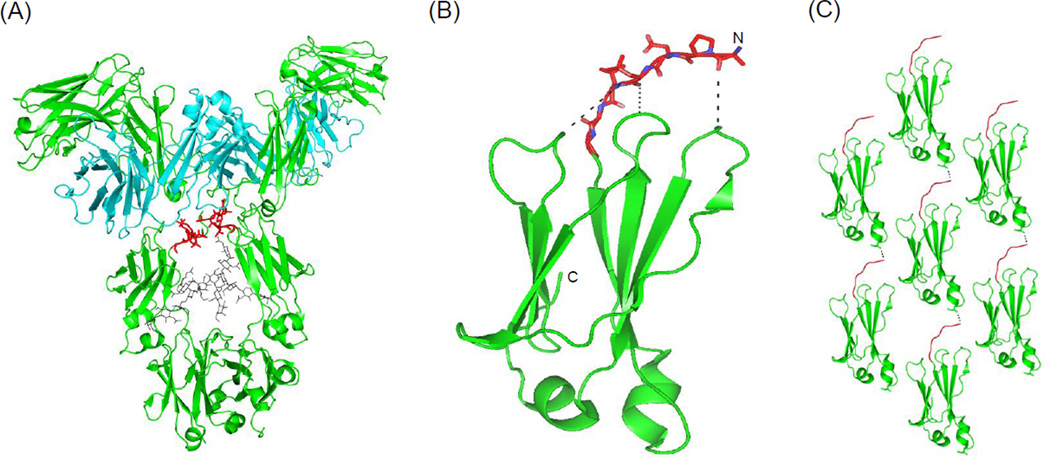
Isolated human immunoglobulin G (IgG) CH2 domains are promising scaffolds for novel candidate therapeutics. Unlike other human IgG domains, CH2 is not involved in strong interchain interactions, and isolated CH2 is relatively stable. However, isolated single CH2 is prone to aggregation. In native IgG and Fc molecules, the N-terminal residues of CH2 from the two heavy chains interact with each other and form hinge regions. By contrast, the N-terminal residues are highly disordered in isolated CH2. We have hypothesized that the removal of the CH2 N-terminal residues may not only increase its stability but also its aggregation resistance. To test this hypothesis we constructed a shortened variant of IgG1 CH2 (CH2s) where the first seven residues of the N-terminus were deleted. We found that the thermal stability of CH2s was increased by 5 °C compared to CH2. Importantly, we demonstrated that CH2s is significantly less prone to aggregation than CH2 as measured by Thioflavin T (ThT) fluorescence, turbidity, and light scattering. We also found that the CH2s exhibited pH-dependent binding to a soluble single-chain human neonatal Fc receptor (shFcRn) which was significantly stronger than the very weak binding of CH2 to shFcRn as measured by flow cytometry. Computer modeling suggested a possible mode of CH2 aggregation involving its N-terminal residues. Therefore, deletion of the N-terminal residues could increase drugability of CH2-based therapeutic candidates. This strategy to increase stability and aggregation resistance could also be applicable to other Ig-related proteins.
Figures
References
-
- Waldmann TA. Immunotherapy: past, present and future. Nat Med. 2003;9(3):269–277. - PubMed
-
- Casadevall A, Dadachova E, Pirofski LA. Passive antibody therapy for infectious diseases. Nature reviews Microbiology. 2004;2(9):695–703. - PubMed
-
- Schrama D, Reisfeld RA, Becker JC. Antibody targeted drugs as cancer therapeutics. Nature reviews Drug discovery. 2006;5(2):147–159. - PubMed
-
- Carter PJ. Potent antibody therapeutics by design. Nat Rev Immunol. 2006;6(5):343–357. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
