

Temporal changes of cytochrome P450 (Cyp) and eicosanoid-related gene expression in the rat brain after traumatic brain injury
- PMID: 23642095
- PMCID: PMC3658912
- DOI: 10.1186/1471-2164-14-303
Temporal changes of cytochrome P450 (Cyp) and eicosanoid-related gene expression in the rat brain after traumatic brain injury
Abstract
Background: Traumatic brain injury (TBI) induces arachidonic acid (ArA) release from cell membranes. ArA metabolites form a class of over 50 bioactive eicosanoids that can induce both adaptive and/or maladaptive brain responses. The dynamic metabolism of ArA to eicosanoids, and how they affect the injured brain, is poorly understood due to their diverse activities, trace levels, and short half-lives. The eicosanoids produced in the brain postinjury depend upon the enzymes present locally at any given time. Eicosanoids are synthesized by heme-containing enzymes, including cyclooxygenases, lipoxygenases, and arachidonate monoxygenases. The latter comprise a subset of the cytochrome P450 "Cyp" gene family that metabolize fatty acids, steroids, as well as endogenous and exogenous toxicants. However, for many of these genes neither baseline neuroanatomical nor injury-related temporal expression have been studied in the brain.In a rat model of parietal cortex TBI, Cyp and eicosanoid-related mRNA levels were determined at 6 h, 24 h, 3d, and 7d postinjury in parietal cortex and hippocampus, where dynamic changes in eicosanoids have been observed. Quantitative real-time polymerase chain reaction with low density arrays were used to assay 62 rat Cyps, 37 of which metabolize ArA or other unsaturated fatty acids; 16 eicosanoid-related enzymes that metabolize ArA or its metabolites; 8 eicosanoid receptors; 5 other inflammatory- and recovery-related genes, plus 2 mouse Cyps as negative controls and 3 highly expressed "housekeeping" genes.
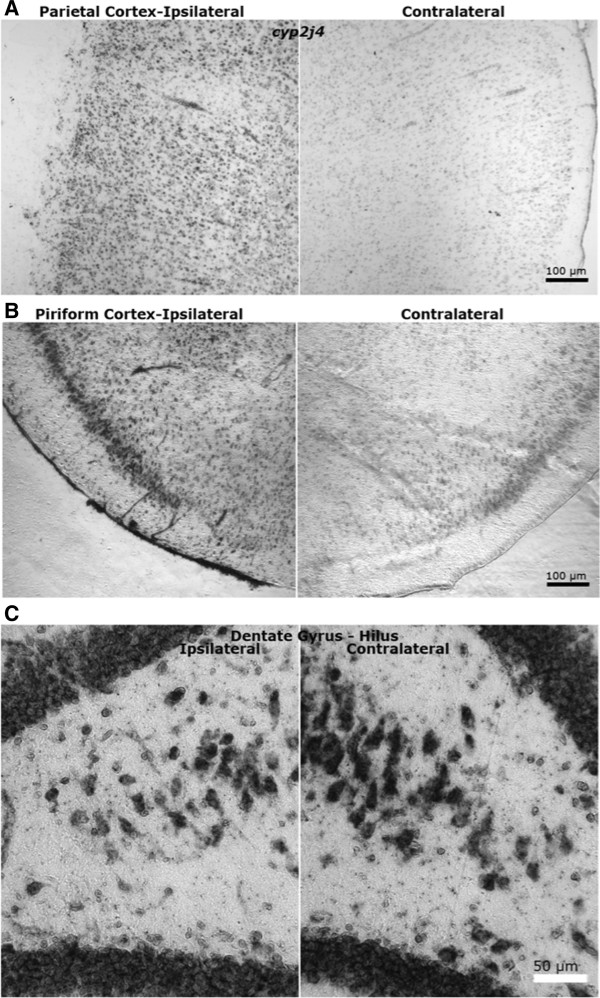
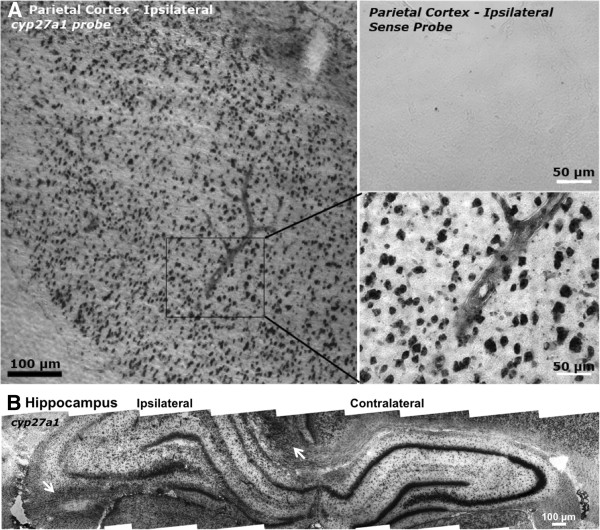
Results: Sixteen arachidonate monoxygenases, 17 eicosanoid-related genes, and 12 other Cyps were regulated in the brain postinjury (p < 0.05, Tukey HSD). Discrete tissue levels and distinct postinjury temporal patterns of gene expression were observed in hippocampus and parietal cortex.
Conclusions: The results suggest complex regulation of ArA and other lipid metabolism after TBI. Due to the temporal nature of brain injury-induced Cyp gene induction, manipulation of each gene (or its products) at a given time after TBI will be required to assess their contributions to secondary injury and/or recovery. Moreover, a better understanding of brain region localization and cell type-specific expression may be necessary to deduce the role of these eicosanoid-related genes in the healthy and injured brain.
Figures
References
-
- Nelson DR, Zeldin DC, Hoffman SM, Maltais LJ, Wain HM, Nebert DW. Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics. 2004;14(1):1–18. doi: 10.1097/00008571-200401000-00001. - DOI - PubMed
-
- Kozak W, Kluger MJ, Kozak A, Wachulec M, Dokladny K. Role of cytochrome P-450 in endogenous antipyresis. Am J Physiol Regul Integr Comp Physiol. 2000;279(2):R455–R460. - PubMed
-
- McGiff JC, Carroll MA. Cytochrome P450-dependent arachidonate metabolites, renal function and blood pressure regulation. Adv Prostaglandin Thromboxane Leukot Res. 1991;21B:675–682. - PubMed
-
- Makita K, Falck JR, Capdevila JH. Cytochrome P450, the arachidonic acid cascade, and hypertension: new vistas for an old enzyme system. FASEB J. 1996;10(13):1456–1463. - PubMed
-
- Fitzpatrick FA, Murphy RC. Cytochrome P-450 metabolism of arachidonic acid: formation and biological actions of "epoxygenase"-derived eicosanoids. Pharmacol Rev. 1988;40(4):229–241. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
