

The effect of hydration state and energy balance on innate immunity of a desert reptile
- PMID: 23642164
- PMCID: PMC3660207
- DOI: 10.1186/1742-9994-10-23
The effect of hydration state and energy balance on innate immunity of a desert reptile
Abstract
Introduction: Immune function is a vital physiological process that is often suppressed during times of resource scarcity due to investments in other physiological systems. While energy is the typical currency that has been examined in such trade-offs, limitations of other resources may similarly lead to trade-offs that affect immune function. Specifically, water is a critical resource with profound implications for organismal ecology, yet its availability can fluctuate at local, regional, and even global levels. Despite this, the effect of osmotic state on immune function has received little attention.
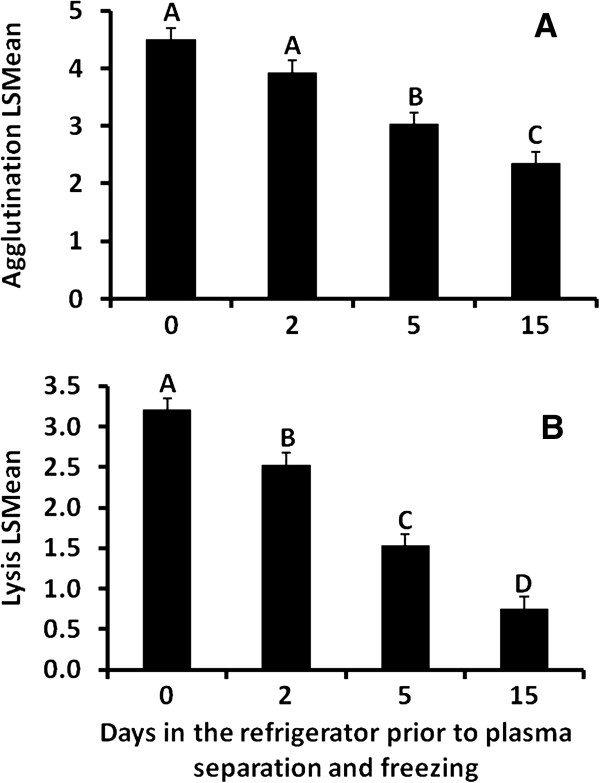
Results: Using agglutination and lysis assays as measures of an organism's plasma concentration of natural antibodies and capacity for foreign cell destruction, respectively, we tested the independent effects of osmotic state, digestive state, and energy balance on innate immune function in free-ranging and laboratory populations of the Gila monster, Heloderma suspectum. This desert-dwelling lizard experiences dehydration and energy resource fluctuations on a seasonal basis. Dehydration was expected to decrease innate immune function, yet we found that dehydration increased lysis and agglutination abilities in both lab and field studies, a relationship that was not simply an effect of an increased concentration of immune molecules. Laboratory-based differences in digestive state were not associated with lysis or agglutination metrics, although in our field population, a loss of fat stores was correlated with an increase in lysis.
Conclusions: Depending on the life history of an organism, osmotic state may have a greater influence on immune function than energy availability. Thus, consideration of osmotic state as a factor influencing immune function will likely improve our understanding of ecoimmunology and the disease dynamics of a wide range of species.
Figures
References
-
- Lochmiller RL, Deerenberg C. Trade-offs in evolutionary immunology: just what is the cost of immunity? Oikos. 2000;88:87–98. doi: 10.1034/j.1600-0706.2000.880110.x. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
